
Numerous small marine shells (smaller than 11mm) have been found at the site. These remains are not burnt, unlike the great majority of mussels with which they were mixed in the ditch of the enclosure and in the pit (Figure 9). These shells do not seem to have been consumed by past humans (see section 4.1). In order to understand the origin of these shell fragments, they have been compared with other remains from modern mussel-beds. Bronze Age populations could have accidentally transported these small shell remains fixed to and mixed with collected mussels. Mussels originating from four modern-day coastal areas (Sangette, Nord-Pas-de-Calais; Saint-Honorine-des-Pertes, Basse-Normandie; Quiberon, Brittany and Lérat, Pays-de-La-Loire) located at the Channel and Atlantic coasts have been collected. These mussels were cleaned in a strainer and associated shell remains compared with archaeological remains. Fragment sizes in modern mussels vary (between 2 and 20mm) and these fragments show rounded outlines caused by marine erosion. By contrast, the sizes of archaeological remains identified at Clos des Châtaigniers are more uniform (between 5 and 10mm), and show angular and regular breaks.
Identified shell species (Barnea candida, Donax sp., Macoma balthica, Spisula sp. and Abra sp.) and those fragmented shells measuring between 5 and 10mm could correspond to residues from the stomach contents of fish. The fish would have been gutted and the entrails discarded by the inhabitants, together with any shellfish remains. Several fish species eaten by humans have a diet that includes shellfish, such as European eel (Anguilla anguilla), conger (Conger conger), plaice (Glyptocephalus cynoglossus), Ballan wrasse (Labrus bergylta), haddock (Melanogrammus aeglefinus), hake (Merluccius merluccius), sole (Solea solea), gilthead bream (Sparus aurata), turbot (Psetta maxima), pollack (Pollachius pollachius) or dogfish (Scyliorhinus sp.) (Figure 10) (Muus et al. 2005; Teletchea 2009). One should bear in mind that the presence of such remains could also result from the accumulation of dead aquatic birds or from faecal waste (Erlandson and Moss 2001; Van Leeuwen 2012). Thus, shell remains could also originate from animal stomach contents. It is important to note here that remains of three unidentified fish and one bird have been discovered at Clos des Châtaigniers.
| Latin name | Common name | Marine shell consumed |
|---|---|---|
| Anguilla anguilla | European eel | + |
| Conger conger | European conger | + |
| Cyclopterus lumpus | Lumpfish | + |
| Diplodus sargus | Sargo | + |
| Diplodus vulgaris | + | |
| Gadus morhua | Atlantic cod | + |
| Glyptocephalus cynoglossus | Plaice | bivalves |
| Hypotremata sp. | Ray | mostly mussels |
| Labrus bergylta | Ballan wrasse | + |
| Limanda limanda | Common dab | gastropods and bivalves |
| Melanogrammus aeglefinus | Haddock | + |
| Merluccius merluccius | European hake | + |
| Microstomus kitt | Lemon sole | mussels |
| Molva molva | Common ling | + |
| Pagellus erythrinus | Common pandora | + |
| Pagrus pagrus | Red porgy | + |
| Pegusa lascaris | Sand sole | + |
| Platychthys flesus | European flounder | mussels |
| Pleuronectes platessa | European plaice | bivalves with thin shells |
| Pollachius pollachius | Pollack | + |
| Psetta maxima | Turbot | bivalves |
| Scyliorhinus canicula | Small-spotted catshark | + |
| Scyliorhinus stellaris | Nursehound | + |
| Scorpaena scrofa | Red scorpionfish | + |
| Solea solea | Common sole | bivalves with thin shells |
| Sparus aurata | Gilt-head bream | mostly bivalves |
| Trisopterus luscus | Pouting | small bivalves |
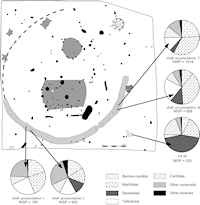
Analysis of bivalves other than mussels allowed us to understand the distribution of these small shells within the settlement (Figure 11). Pit 28 is unique, with a strong presence of Semelidae (44% of the NISP) while Tellinacea is almost absent (3% of the NISP). The distribution of Tellinacea, Pholadidae, Mactridae and Cardiidae is uniform in the ditch of the enclosure. By contrast, Semelidae are totally absent from accumulation 1. This distribution could potentially correspond to the preparation of fish or birds that have different diets.
| Order | Family | Genus | Species |
|---|---|---|---|
| Veneroida | Pholadidae | Barnea | candida |
| Mactridae | Spisula | sp. | Mactra | sp. |
| Semelidae | Abra | sp. | Scrobicularia | plana |
| Tellinacea | Donax | sp. | Macoma | balthica |
| Cardiidae | Cerastoderma | sp. | Acanthocardia | sp. |
Theodoropoulou (2007, 178) points out that, on some archaeological sites, small-sized marine shells could originate from stomach contents of mammals and aquatic birds or fish. However, she highlights the fact that it is very difficult to establish such an origin given the lack of reference studies. Van Neer and Pieters (1997) mentioned that, in a 15th-century coastal village in Belgium, remains of plaice (Pleuronectes platessa, 130 individuals) have been discovered in association with bean clam shells (Donax vittatus), interpreted as the stomach contents of these fish. Likewise, in waste areas of the medieval castle of Boves (Somme), all the identified marine shells (8 at species level and 9 at genus level) originate from the preparation of fish used in human diet (Dupont 2005).
In order to answer this type of interrogation more precisely, it would be interesting to propose some criteria for the identification of aquatic animals from their stomach contents, through the development of studies and bibliographical research on the diet of these faunas. The results could demonstrate the consumption of other aquatic animals, whose skeletal remains were not preserved, marine shells being the only indicators of their initial presence.