
Fig. 2: Fragment of right brow tine Megaceros (?ssp.) giganteus.
Preliminary taxonomic classifications of the cervid material were published by Mania (1991b); additional taxonomic observations were made by the author in the course of the archaeological investigation of the antler material. The crown forming Cervus elaphus (Red Deer) dominates the antler material, while only a few pieces of antler belong to other cervids. Four antler fragments (150/216; 187/150; 262/20; 76/19) belong to Capreolus sp (Roe Deer). Some antler bases (106/299; 106/300; 118/41; 14/37; 149/151; 153/B3; 176/60; 186/39; 198/66; 198/70; 437/2; 438/13; 77/B3; 83/B2) show features that could relate this material to Dama sp. (Fallow Deer), but do not form conclusive proof of the existence of this species (H.-D. Kahlke, Weimar, personal comment). There is, however, no doubt about the find of a right brow tine (172/282) of Megaloceros (?ssp.) giganteus (Giant Deer) (Fig. 2; classification by H.-D. Kahlke) previously unknown in Bilzingsleben (survey of the Bilzingsleben faunas in Mania 1991b).
Fig. 2: Fragment of right brow tine Megaceros (?ssp.) giganteus.
2820 antler finds were registered during the excavations of 1969-1993. The intense fragmentation of the material made a detailed morphological description of each find necessary. A descriptive system was developed, which subdivided an antler into some 80 elements and element fragments, each with a specific code number. (Fig. 3a and Fig 3b, and Table 1).
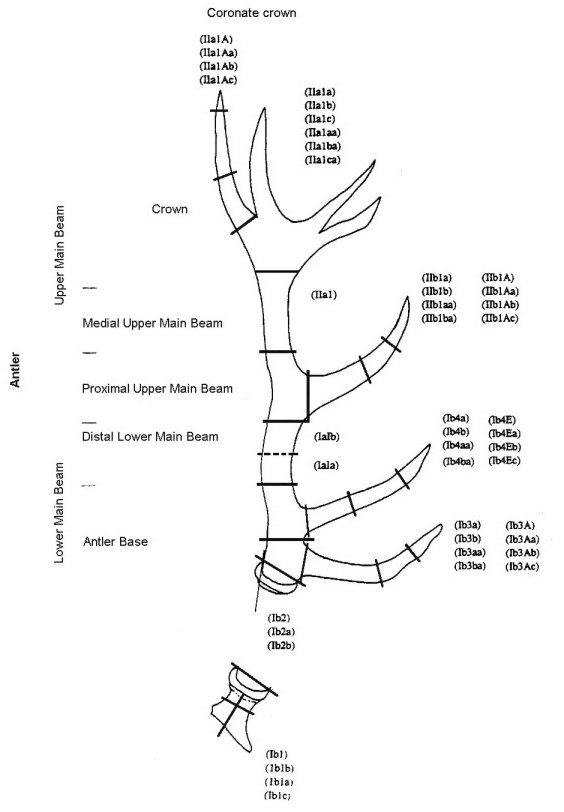
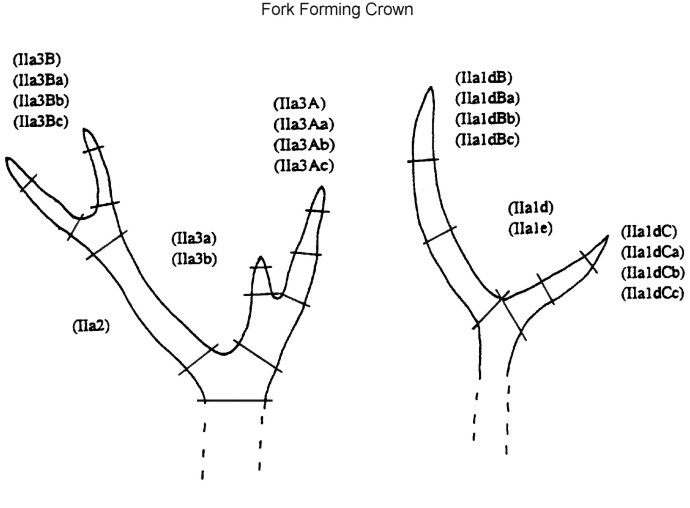
Fig. 3a (left) and Fig 3b (right): The parts of a red deer antler. Explanation of code numbers as used in the main
antler database can be found in Table 1).
On the basis of this subdivision, each antler find can be described as an antler element or as a part of it. In this way large antler finds can be described quite precisely, but small finds must often be described in more general terms. Due to the variable forms of red deer antler crowns, the assemblage (corresponding to the code numbers of the descriptive system in Table 1) includes a relatively high number of crown elements. For example, crown forks are only distinguishable from their particular position within the antler, so isolated finds of crown tines or crown tine fragments can not be determined accurately using the detailed descriptive system. General terms are therefore also included to cover this e.g. 'crown tine' (see Table 1).
The fragmentary character of single finds can be shown by a general morphological description of the antler material (Table 2). Tines form by far the dominant part (66.8%) of the material, whilst 12.5% consists of antler cortex fragments, and 20.6% are antler beams and antler beam fragments. 73.8% of the latter come from the lower and 20.1% from the upper beam, while the remainder of the beam material cannot be identified. The first observation of note is that fragments from the lower beam are more frequently represented than parts from the upper beam. A more exact characterisation of the finds is given by their detailed morphological classification in the morphology dataset. Counts of individual elements shed further light on the disproportionate representation of lower and upper beam elements (Fig. 4).
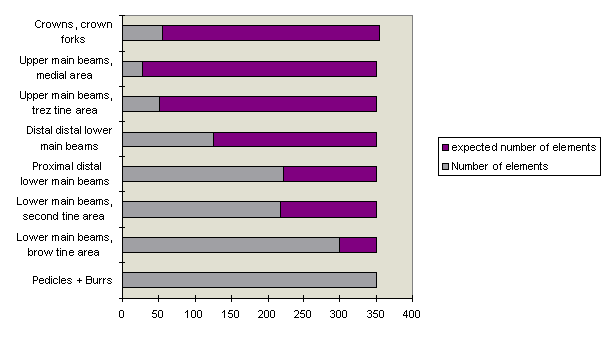
Fig. 4: Ratio of expected and found beam elements.
From about the trez tine upward, the number of fragments becomes more or less constant. Comparison of the number of beam fragments forming part of element 'sums' (antler finds made up of 2 or more elements) with those of the isolated fragments (Table 3) demonstrates consistently low values for the isolated ones. It may be concluded that the most frequent parts from the lower beams (e.g. from the antler base and lower main beam) occur mainly in element sums.
| Element sum | isolated | |
|---|---|---|
| Crowns, crown forks | 0.07% | 4.3% |
| Medial upper main beams | 0.07% | 2% |
| Upper main beams, trez tine area | 0.7% | 3.1% |
| distal distal lower main beams | 6.8% | 2.5% |
| proximal distal lower main beams | 12.7% | 3.7% |
| Lower main beams, second tine area | 14.4% | 2% |
| Lower main beams, brow tine area | 20.1% | 2.2% |
| Pedicles + burrs | 21.7% | 4.3% |
Another interesting observation can be made from the detailed morphological description : antler bases and lower parts of beams (of both shed and unshed antlers) show variable forms, for example with regard to the preserved brow and trez tine parts. Finally, the question of matching beam elements with the abundant tine material had to be examined in both a practical and a theoretical way. Practically this was done by looking for matching breaks, while the other way was to examine the antler material using the detailed morphological descriptions. The count of recovered proximal fractured tines should theoretically be equivalent to the number of distal fractures. Comparison of the actual number (476) of proximal fractured tines with the number of distal fractures shows that the latter are in fact under-represented (Fig. 5) and that, therefore, the antlers represented by the excavated material are incomplete.
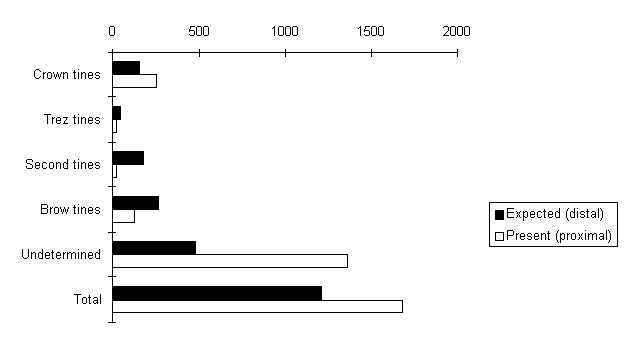
Fig. 5: Ratio of expected and found tine elements.
The abundance of isolated tines does not correspond with the number of tines needed for the beams found, nor does the number of lower and upper beams agree. The process that produced the assemblage can not therefore have consisted of a series of simple activities such as the breaking up of complete antlers and of the subsequent preservation of all resulting antler fragments in the excavated area. A more complex history of accumulation or loss of antler fragments has to be considered.
© Internet Archaeology
URL: http://intarch.ac.uk/journal/issue8/vollbrecht/en/4.html
Last updated: Thu Aug 10 2000