
Cite this as: Winder, J.M. 2015, Oysters and Other Marine Shells, in M. Atkinson and S.J. Preston Heybridge: A Late Iron Age and Roman Settlement, Excavations at Elms Farm 1993-5, Internet Archaeology 40. http://dx.doi.org/10.11141/ia.40.1.winder
Oyster and other marine mollusc shells were examined to determine how significant the exploitation of marine and riverine resources had been to the economy of the settlement, and to establish the importance of the river as a communication route for the inhabitants. Just over 109kg of marine molluscs were recovered from a total of 108 contexts located within 75 features. These derived from all parts of the site except Areas D and R, with the majority coming from central and southern zones of the settlement.
The shell was recovered from the site by hand retrieval and through wet sieving of bulk samples. Where large dumps of shell were present, samples were taken and an estimate made of the percentage of shell removed. The shell was then carefully washed and species identified and counted. Oysters (Ostrea edulis L.) were sorted into left and right valves, measurable and unmeasurable shells. The criteria for selection for measurement were the presence of the hinge or ligament scar, the adductor muscle scar, and at least two-thirds of the shell intact. The numbers of oyster valves and other marine mollusc shells were recorded by context and by feature. Abundance by phase was also calculated.
The criterion for inclusion of a sample for analysis was a minimum number of at least thirty measurable individuals of either left or right valves. Measurements of length and width in millimetres were recorded for oysters from nine contexts in which an adequate number of shells remained in relatively good condition. The oyster shells from five of these were selected for more detailed examination. Evidence for infesting or encrusting epibiont organisms and various descriptive characters were recorded on a presence or absence basis for each shell. The methods are described in full in Winder (1992). The percentage of shells in each sample affected by each type of infestation was calculated, as was the frequency of descriptive characters.
Comparisons of size and size distribution in oyster samples were carried out by the parametric two sample t-test (Blalock 1972, 219-41; Bailey 1959, 33-42) and the non-parametric Mann Whitney test (Blalock 1972, 255-62; Bailey 1959, 153-60) on an intrasite basis between the oyster samples from the nine suitable contexts. These samples were also compared on an intersite basis with those from other Roman sites and from modern oyster beds.
The percentage frequencies of the various types of infestation evidence in each sample were used to make comparisons between the Elms Farm oysters and samples of Roman oysters from elsewhere in Britain using Principal Component Analysis (PCA) (Morrison 1967; Johnston 1978; Shennan 1988).
The descriptive features of the shells are collated as an aid in determining whether the oysters have been farmed.
A list of the contexts from which oysters and other marine molluscs were recovered is given in Table 153 (.csv). This provides the contexts containing shells in numerical order, the feature number, feature type, number of shells, and comments on wear.
Table 154 (.csv) provides the same information as Table 153 (.csv) but arranged by feature number. Table 155 (.csv) presents the percentage frequencies of the different species of shell (as a total of all contexts in each feature).
From the marine molluscs recovered, one hundred and three contexts from sixty-one features were finally selected for analysis (the remainder were considered to have been possible contamination from earlier or later deposits).
Both the spatial distribution across the site and the the proportional representation of shell in the different localities has been considered. The total weight of recovered oyster shell was 291,483g. Oyster shells were recovered from Excavation Areas E, H, I, J, K, L, M, N, P and W. The distribution by weight in these areas varied. Areas E, W, P, H and L each contributed less than 5% of the total shell. Most shell was recovered from Area I (57.9%), with 14.6% from Area J, 9.9% from Area N, 9.6% from Area M and 5.3% from Area K (Table 156 and Table 157 (.csv)).
Of the areas from which most shell was recovered, Areas I and J are in the perceived centre of settlement, each with different character, though possibly associated functions. Areas N, M and K are in the Southern Zone with its regular occupation plots dominated by pitting. Furthermore, Areas I, J and K are clustered not only along the east-west route of Roads 2 and 3 that presumably run from a crossing of the River Chelmer but also around the temple complex.
Of the areas yielding few shells, Area E is in the Northern Zone, which possibly represents a mix of low-level occupation and animal husbandry within its plots. Area W is in the Hinterland Zone, essentially the immediate agricultural landscape. Areas P and L are in the pit-dominated Southern Zone. Area H is in the centre of settlement where each area has a different, though possibly inter-related, character.
The percentage distribution of oyster shell by weight across the site therefore shows a concentration in the south-west of the site, near the junctions of Roads 2, 3 and 4 in what is the perceived Central Zone of the Roman settlement. There is also a concentration in the Southern Zone. It should be noted that very few shells were recovered from Area H, which is designated as a potential market area that had evidently been cleared and surfaced.
The oyster shell groups selected for analysis were all retrieved from pits that were located in Areas H, J, K, M and N.
Table 158 and Table 159 (.csv) show how the contexts containing oyster shell were distributed through time. The greatest weight of oyster shell (56.1%) was recovered from contexts attributed to Period 3B (Early Roman), with 16.2% in Period 4 (mid-Roman) contexts, 9.6% in contexts assigned to Period 4-5, and 9.3% in Period 5 (Late Roman) contexts. Each of the other phases contributed less than 4% of shell by weight. Not only is there an apparent decrease in abundance through time but the area of the site over which oyster shells are recovered contracts through time. While in Period 3 twenty-eight contexts with shell are scattered over six areas, by Period 6 (Early Saxon) ten contexts with shell are distributed over just three areas.
The oyster shells that were measured and examined in detail were from Periods 3-6.
A total of 6637 oyster shells (Ostrea edulis L.) were recorded. The actual number of oysters present on site was greater than this because some samples represent just a fraction of the deposit. There were also sixty-nine examples of Buccinum undatum L. (whelks), fourteen Tapes sp (carpet shell valves), ninety-six Mytlius edulis L. (mussel valves), seven Cerastoderma edule (L.) (cockle valves) and three Littorina littorea (L.) (periwinkles).
Mussels were mostly found in Period 3 deposits (seventy-six shells, 80% of total) in the four areas E, I, K and L. Very small numbers were found in Period 4 and 5 deposits (4% and 3% respectively), also in Area K. Five per cent of mussels were retrieved from Period 4-5 in Area N. Mussels commonly occur from high in the intertidal zone to depths of a few fathoms attached by byssus threads to rocks and other hard objects within sheltered harbours and estuaries as well as rocky shores of the open coast (Tebble 1966). They are predominantly estuarine and large commercial beds are all situated in areas subject to reduced salinity (Graham 1956). In suitable estuaries, where there are large areas of stony or gravelly bottom exposed between tide marks, mussels may form very large banks or scars. Some beds are never uncovered by the tide but these rarely extend beyond the immediate sublittoral zone. Mussels can be gathered by hand, by raking in submerged pools and channels, and by dredging. However, the silty nature of the Blackwater estuary makes it a relatively poor habitat for mussels, which may explain their relative sparcity in excavated contexts.
Whelks were mostly found in deposits dating to Periods 4 and 4-5 (6% and 57% respectively) in Areas N, M and J. Only 1.4% of whelks belonged to Period 3 in Area I, while 10% of whelks were attributed to Period 5 in Areas K and M. Whelks live in the low inter-tidal zone down to deep water in estuaries and open seas on a variety of sea bottoms but with a preference for mud mixed with sand and shells. They are found all round our coasts, but whelk fishing is only locally important today on the east and south coasts. In the sea the whelk meat grows white and fills the shell while in estuaries and rivers the flesh is darker and may be poor quality (Hancock 1967). Whelks are usually caught using baited pots but sometimes in east coast estuaries by 'trotting', where baited lines are laid across the tide for about six hours and then hauled in.
At the assessment stage eleven contexts were thought to contain enough measurable shells to enable the samples to be used in comparisons of size. These contexts were 4801, 4844, 5144, 5214, 5393, 10891, 11139, 13813, 15152, 15515 and 16213. The numbers of oyster shells in the nine contexts selected for size comparisons are given in Table 160 (.csv). However, contexts 13813 and 16213 actually had too few measurable shells and were therefore excluded from further analyses. Descriptive size statistics for the nine selected oyster samples for left valve maximum width (LVMW), left valve maximum length (LVML), right valve maximum width (RVMW) and right valve maximum length (RVML) are given in Table 161 (.csv).
Taking the left valve maximum width, mean size varied from 77.5mm to 86.2mm with standard deviations between 7.7 and 8.6mm. The smallest oyster shell was 56mm and the largest 105mm in width. The greatest range in size was 48mm in context 10891, and the smallest range was 35mm in context 4844.
It should be noted that the range of sizes in the samples varies with time. The Early Roman (Period 3B) range is 39mm, mid-Roman (Period 4) is 39.8mm, mid-to Late Roman (Period 4-5) is 47.5mm, Late Roman (Period 5) is 43mm, and latest Roman/Early Saxon (Period 6) is 38mm. This indicates a greater range of sizes in samples from mid- to Late Roman (Period 4-5) and Late Roman (Period 5) than from the other periods. This greater range in size is associated with a rise in average size in the mid- to Late Roman (Period 4-5) from 80-84mm followed by a decrease in average size (84-75mm) in the Late Roman phase (Period 5). This may be due to the inclusion of larger 'pied du cheval' type oysters.
Reference to Figure 680 (.xlsx) of intrasite comparisons of size in samples from selected contexts shows that these size differences are statistically significant. This is discussed further under Intrasite comparisons. The implications of a decrease in average size and greater range of sizes might be less selectivity in collection, over-fishing causing a reduction in the numbers of oysters in the preferred size range, or a change of collection method.
Matrices of the results of the intrasite comparisons of size by two sample t-test (assuming unequal variances) and Mann Whitney test, using left valve maximum widths are presented in context order in Figure 678 (.xlsx), by area in Figure 679 (.xlsx), and by phase in Figure 680 (.xlsx).
There is no significant difference between contexts 4801 (Period 4), 4844 (Period 4), 5144 (Period 3B) and 5214 (Period 6) and between most of these and contexts 15152 (Period 5) (bar 5144) and 15515 (Period 4). In this group of samples the mean left valve maximum width (LVMW) measurements are all 80mm or less.
Context 5393 (Period 4) does not significantly differ from context 10891 (Periods 4-5). Context 11139 (Periods 4-5) is not significantly different from context 15515 (Period 4). In this group of samples the mean LVMW measurements are all greater than 80mm.
Figure 679 (.xlsx), with comparisons of size of samples arranged by area of site, shows that the samples within each area are not uniform in size and size distribution. Some samples from the same area are significantly different in size and others are not. Similarly, between different areas of the site, there are generally no clearly indicated similarities or differences. The samples in Areas K and N, which are all significantly different in size from each other, form the exception.
Figure 680 (.xlsx), with comparisons of size of samples arranged by phase, shows that the samples within each phase may be significantly different from each other. Between phases there are also significant differences that reflect the changes in mean size at each phase - as discussed above in discussion of size. The Early Roman sample is significantly different from 25% of mid-Roman samples, 50% of mid- to Late Roman samples, and 100% of Late Roman samples. The mid-Roman samples are significantly different in 75% of comparisons with mid- to Late Roman samples, 50% of Late Roman samples and 25% of latest Roman to Early Saxon samples. Mid- to Late Roman samples are significantly different in all comparisons with Late Roman and latest Roman to Early Saxon samples.
These tests therefore indicate that there is a significant increase in size, and range of size, in oysters from Early Roman to mid- to Late Roman and a subsequent decrease in size and range through to the latest Roman/Early Saxon phase.
Comparisons were made of Elms Farm oysters with other Roman sites. The sizes of shells in the nine selected samples were compared with thirty-seven samples deriving from thirteen other Roman sites, mostly in southern England. Comparisons were made by two sample t- and Mann Whitney tests. These sites include Shapwick (Winder 2000a), Greyhound Yard (Winder 1993a), Alington Avenue (Winder 2002), Halstock Roman Villa (Winder 1993b) in Dorset; The Brooks (Winder 2000b) and Owslebury (Winder 1988) near Winchester in Hampshire; Newport Roman Villa (Winder 1989a) on the Isle of Wight; Pudding Lane (Winder 1984, 1985a) in London; North Shoebury (Winder 1993c) and Colchester (Winder 1993c) in Essex; Tort Hill (Winder 1998b) in Cambridgeshire and the Shires (Monckton and Winder 1992) in Leicester. The size data for all these Roman sites is summarised in Table 162 (.csv).
The matrices of results from these tests are shown in Figure 681 (context order; .xlsx), Figure 682 (phase order; .xlsx) and Figure 683 (area order; .xlsx). The results of these comparisons are difficult to interpret. The results have been divided into groups depending on the ratio of significantly different sample comparisons to non-significantly different comparisons.The first group comprises results of comparisons between Elms Farm oyster samples and other Roman samples between which there is a statistically significant difference in size for all samples. The last group comprises results of comparisons where there is no statistically significant difference in size for seven out of the nine Elms Farm samples. Whether the Elms Farm oysters are considered by context, phase or area, there appears to be no clear pattern of similarity or difference based on samples from different geographical locations. Many Elms Farm oysters show no significant difference from south coast or other Essex oysters.
The Elms Farm oysters do not significantly differ in size and size distribution from many of the other samples. This means that the null hypothesis, that the two samples are derived from the same population, cannot be rejected. It is possible that contexts 4801 (Group 4016), 5144 (Group 430) and possibly 15515 (Group 696) share common size characteristics that are distinct from those of contexts 5393 (Group 432) and 10891 (Group 676). For example, contexts 5393 and 10891 are both significantly different from all the North Shoebury and Colchester samples while contexts 15515 and 5144 show no significant difference from five out of six of the same set of samples; and context 4801 is not significantly different from three of the six samples.
Table 163 gives a summary of the size data relating to samples from modern oyster populations. Figure 684 (.xlsx) shows that in comparison with oyster samples from locations in the Rivers Roach and Colne (MAFF 1960s) - no Blackwater data is available - and from the Colchester Oyster Feast (Colchester Museum 1971), the Elms Farm oysters from contexts 5144 (Group 430), 5393 (Group 432, 10891 (Group 676), 11139 (Group 676), 15515 (Group 696) were all significantly different from the modern samples. Contexts 4801 (Group 4016), 4844 (Group 4016) and 5214 (Group 442) were not significantly different from the modern relaid East coast samples and the modern wild oysters from the south coast in Poole Bay. The modern Colne, Roach and Colchester samples were all native oysters. The modern Poole Bay oysters were natural and unsorted except by dredge bag mesh size. The mean size of the Solent oysters was smaller and the Poole Harbour oysters were larger than the Elms Farm contexts 4801, 4844 and 5214.
Infestation evidence was recorded in only those five contexts where the shells were considered to be in better condition. These were 4801 (Group 4016), 5144 (Group 430), 5393 (Group 432), 10891 (Group 676) and 15515 (Group 696). Evidence was recorded of the burrows of the marine polychaete worm Polydora ciliata, borings of the sponge Cliona celata, boreholes made by gastropod molluscs such as Ocenebra erinacea and Nucella lapillus, and encrusting barnacles, Bryozoa and sand tubes made by Sabellid worms. Full details of these organisms can be found in Winder 1993c.
Table 164 (.csv) presents the infestation data as percentage frequency of evidence for epibiont infestation or encrustation recorded in the samples of oyster shell from Elms Farm and other Roman sites. Principal component analysis was used to compare Elms Farm oysters with all available Roman sites (Figure 195), and with other nearby Roman sites at Colchester and North Shoebury (Figure 196).
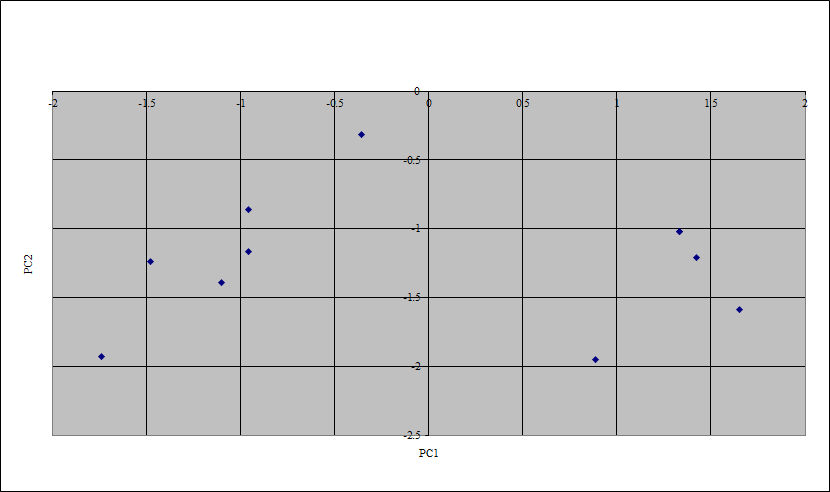
Principal Component Analysis of infestation was undertaken on the Elms Farm oysters and samples of Roman oysters from elsewhere (also see archive). This showed that the Elms Farm oysters share the infestation characters common with other east coast oysters but on a local scale show differentiation from nearby sites. In comparison with the Elms Farm samples, the Colchester oysters have a fairly high percentage of Polydora ciliata (68.6%) and slightly higher levels of Cliona celata (2.7%) but lower levels of barnacles (0.3%), and Bryozoa (0.9%). On the other hand, North Shoebury samples are characterised by high percentages of Polydora ciliata (up to 89.6%), high levels of barnacles (up to 25.8%) and Bryozoa (up to 15.1%), with low levels of Cliona celata (maximum 1.4%) and the presence of sand tubes in one sample (Shoe 299). Sand tubes were also noted in the Elms Farm samples (0.9% in 10891) but all differed from oysters at the other two sites in the presence of boreholes (0.6% to 3.9%). Clearly, the individual combinations of infestation evidence indicate different habitat characteristics of different oyster beds.
Twelve descriptive characters were recorded for oyster shells in the five contexts selected for more detailed recording (4801 (Group 4016), 5144 (Group 430), 5393 (Group 432), 10891 (Group 676) and 15515 (Group 696)). These included relative thickness and weight; chambering and chalky deposits; degree of wear; natural colour or post-burial staining; attachment of adult or spat oysters; irregularity of shape; man-made notches or cuts; and the presence of a ligament. Table 165 (.csv) presents the frequencies with which these characters were recorded in the samples.
Shells from context 4801 show the greatest degree of variation, exhibiting the highest percentages of both thin and thick shells, chambering, wear and flakiness, attached oysters and ligament survival. Shells from context 5393 show much more uniformity in general features with the lowest percentage of thin, thick, heavy and chambered shells, together with few chalky deposits and attached oysters.
Chambering and chalky deposits were recorded in all samples; generally these two features are closely associated - as is the case in four of the samples. In context 4801, however, chalky deposits were less important than chambers. The formation of chambers and chalky deposits is related to a need to reduce, or alter the shape of, the internal volume of the shell cavity between the two valves in response to a sudden reduction in meat volume. The animal is surrounded by an enveloping structure called the mantle which is responsible for shell formation and must remain in contact with the shell at all times. When subjected to rapid changes in salinity (usually an increase) or when depleted after spawning, the animal shrinks. A very thin-walled partition is often laid down internally, very rapidly trapping fluids and sediments, to form a chamber, or a pad of micro-crystalline structure is formed to similarly alter internal shape. Chambers and chalky deposits can therefore be an indicator of either shallow-water conditions (as in creeks and tidal inlets) where evaporation increases salinity, or breeding activity.
All the samples had irregular-shaped shells (ranging from 12.6% in 10891 to 4.9% in 5144). Irregularity in oyster shell shape is a response to growth in restrictive physical conditions such as settlement on a rough substrate and/or overcrowding caused by many oysters competing for space. Shape can therefore be a guide as to whether the oyster sample was derived from a natural population where irregularity may be common, or whether the oysters have been farmed, where they are separated and spread out on the sea bed and thus achieve a more regular shape. It should be noted that the absence of irregularity cannot necessarily be taken to imply that the oysters were relaid since repeated dredging on a natural bed can produce the same effect.
Cut marks were present in low numbers in all but context 4802. Cuts and notches are thought to be caused during the opening of oysters and the removal of meat. Cuts suggest that the oysters were being eaten alive, or at least opened while alive.
The ligament survived in some specimens from each context, with the highest proportion from 4801 which was also characterised by highest levels of wear and flakiness. This probably indicates that the wear has probably been mechanically produced in some samples as well as resulting from the breakdown of the organic component of the shell.
All the samples included shells to which other oysters were attached (2.4% in 5144 to 8.9% in 4801). The presence of small spat, or more mature oysters, attached to an oyster shell is an indication firstly that the population from which it is derived is actively propagating; and secondly that the population is likely to be natural or wild. By contrast, if the oysters had been farmed, the attached spat or young oysters would have been divided or separated off before marketing and relaid for fattening. This does not appear to have happened, suggesting a lack of active management of the oyster-beds.
It is appropriate here to discuss the significance of the marine mollusc remains at the Roman village at Elms Farm near Heybridge in the light of the evidence provided by the oyster and other shells recovered there. The Essex coast is famous for its oysters (Benham 1993). The River Blackwater, and the town of Maldon are especially renowned for their oyster production and trade (Benham 1993; Yonge 1960, 152-3; Philpots 1890, 390-400). Elms Farm is close to these places with their long association with the oyster industry; and is just 400m from a tributary of the River Blackwater, the River Chelmer, to which it is postulated to have been linked by Road 2. The settlement was therefore ideally situated to exploit a nearby abundance of oysters.
It was initially proposed in the interim report that oysters and shellfish could not have held an important role in the diet and economy of the community because there were too few shells recovered from such a wide area occupied over such a long time 'This lack of evidence for coastal trade is part of a wider absence of indicators for the role of marine and riverine resources in the settlement's economy. Most obvious is the rarity of marine molluscs; only half a dozen or so features and deposits yielded more than the occasional shell, while only one excavated feature contained a significant deposit of mussels' (Atkinson and Preston 1998, 108).
This idea needs to be re-examined now that the marine molluscs have been studied in detail. Nearly 7,000 oyster shells were actually excavated from this site. This is an underestimate of the number of shells actually present because excavations did not extend over the whole site, and larger deposits of shells were sub-sampled. In comparison with quantities of oyster shells recovered from thirteen other Roman sites, numbers vary from 10,810 valves at the Greyhound Yard Roman town site in Dorset, to just 383 valves from the Roman road at Tort Hill in Cambridgeshire, so the Elms Farm shells represent a not inconsiderable amount of oysters. In any terms, with 6637 oyster shells, marine molluscs cannot be called 'rare' at this site. Thirty-seven contexts had more than twenty shells, fourteen contexts had more than one hundred shells. Regarding mussel shells, it should be noted that this type of marine mollusc rarely survives intact in archaeological deposits and is usually found in a comminuted state or even as a blue, almost fibrous, layer. All shells are likely to be adversely affected by the harsh acidic gravel conditions (which were noted as a cause of poor survival in fish bones in Atkinson and Preston 1998, 108) to the point where they might be totally destroyed.
The number of shells to be found on a site depends on various factors, apart from soil conditions and post-deposition treatment. The main consideration would be the balance pertaining in the community between production, consumption, processing and trade. Only coastal or estuarine sites may be areas of shellfish production. The settlements may fish oysters for home consumption, in which case all shells could potentially be disposed of on site. The settlement may trade inland all, or just surplus, stocks to the local or wider hinterland so that potentially only a fraction of oysters fished could be disposed of on site. The numbers of shells on site could therefore be small in relation to the quantity of shellfish actually being handled by a community that was heavily dependent economically on shell fishing.
Large deposits may occur in waterfront locations where the community relied to a large extent, maybe seasonally, on shellfish in the diet, as in Mesolithic shell middens (Mellars 1978) and the Ertebolle culture in Denmark (Andersen and Johansen 1986). Huge dumps of shell can result from marine mollusc processing, usually where the meat is removed from the shells and preserved, for example the oyster middens on which the early medieval town of Poole in Dorset is built (Winder 1992), and the large 12th-century shell midden on the south side of Poole Harbour at Ower Farm, generated when cockles were pickled for a nearby monastery (Winder 1991). Waterfront trading of oysters is thought to have been responsible for the substantial dump of oyster shells beneath the Roman open-work Thames-side piers uncovered beneath Pudding Lane in London (Winder 1984, 1985a). These are all special instances and not typical of the quantities of shell usually recovered during excavations whether of coastal or inland sites.
The habit of recycling oyster and other shells may also account for large dumps on sites, or for the absence of large dumps. For example, ditches are often backfilled with shells as in Saxon Hamwic (Winder 1998a), and the Billingsgate excavations in London revealed the way oysters had been used to infill behind waterfront revetments (Brigham 1990). Floor surfaces and pathways have also been constructed mainly from shell (Six Dials: Winder 1985b; 1986; 1997; Corfe Castle: Winder 1989b). Conversely, recycling may involve the destruction of shells so that few shells are evident on site. Examples of this are the use of shells for mortar between stonework - especially in arches, for making lime, poultry grit, shell-tempered pottery, and particularly in Roman times for medicines and cosmetics (Radcliffe 1931; Masefield 1899; Haman 1893; Nealey 1931; Roughly 1923; Swanton 1898). In other cases oyster shells are spread on fields as fertiliser (calcium neutralising acid soils which cause the shells to delaminate and disintegrate), or - most importantly in oyster production areas - they are put back on the sea bed as the preferred substrate for oyster spat settlement.
The point being made here is that the absence of enormous numbers of oyster shells, from a site which is in a prime location for oyster fishing, is not necessarily an indication that there was no fishing industry.
It is relevant at this stage to discuss in general what is known about oyster exploitation in the Roman period. It is quite clear that the arrival of the Romans in Britain created a widespread demand for, and exploitation of, this particular marine resource which had been largely neglected in the preceding Iron Age. It is not known why this should have been the case. It is possible that climatic conditions in the Iron Age were unfavourable for oyster breeding, or maybe oysters were abundant but not a preferred food source. We only know that oyster shells have seldom been excavated from British Iron Age sites (Owslebury in Hampshire being an exception) and Heybridge is no exception. Only 0.09% by weight of oyster shell was recovered from four contexts in the late pre-Roman Iron Age and transition to Early Roman periods (2 and 2-3). However, 56% of the oyster shells for the whole site were retrieved from eleven phase 3B contexts (late 1st to mid-2nd century AD). This sudden influx of oysters, widespread over the site in pits, trenches, spreads and layers, must surely mark the appearance of Romans (or Romanised others) at Heybridge.
Evidence was sought in the shells to distinguish between oysters from wild or natural beds (Winder 1993c: Model 1, 283-5; Model 2, 285-8) as opposed to cultivated or relaid ones (Winder 1993c: Model 4, 289-92). Oysters from natural beds would be expected to have irregular shapes, adhesion between individuals, oyster spat attached, and a wide range of sizes and ages. In intertidal beds a slow growth rate leading, in some instances, to the phenomenon of 'stunting' might also be expected in oysters spending regular periods of time out of the water. A few 'stunters' were recorded in the Elms Farm samples.
Relaying of young oysters from natural beds where they have bred into new places, which enhance the fattening up of the meat, is the first stage in oyster cultivation or farming. Fattening grounds tend to be in enclosed harbours, estuaries and creeks. The characteristics of samples of relaid oysters might be a restricted age and size range because of cropping at a specific age. Warm, shallow nutrient-rich waters may lead to higher levels of infestation. Where muddy substrates have been consolidated with broken shells, twigs and branches prior to relaying, such 'shram' might be incorporated in the developing shells of the relaid oysters. Shape of shells would be less distorted because individual oysters would have been separated. Spat and attached oysters would be rare.
The Heybridge oysters provided only evidence to suggest that they were fished from natural beds either by hand collection in the intertidal zone or by dredging inshore shallow waters.
Populations of wild oysters in Roman Britain would have been easily accessible and plentiful. An examination of nearly 40,000 shells, together with artefacts and structures from thirteen Roman sites in Britain has failed to provide firm evidence for any direct cultivation or farming (i.e. relaying) of oysters. The only indication that some kind of stock improvement may have been attempted is that shells found on the Roman Thames waterfront beneath Pudding Lane in London changed from predominantly irregular shapes with a lot of infestation in the 1st century AD to more regular shaped cleaner ones in the 2nd century AD (Winder 1984; 1985a). However, other explanations for this can be proposed. It might be that supplies were obtained from a different source where shell characteristics were different, or that extensive dredging on the same oyster bed had split up the naturally occurring clumps of oysters and spread out the individual oysters, which were then able to achieve their full growth potential without restrictions.
There is no evidence to prove that marine as well as estuarine shellfish were being exploited at Heybridge. All the marine mollusc species could have grown in either fully marine or brackish conditions. Oysters and mussels both have a preference for reduced salinity. The frequency of long oyster shells and elongated ligament scars suggest growth on relatively softer or muddier grounds. Similarly the occurrence of chambers and chalky deposits may indicate shallower waters where salinity can rapidly change. These features point to shallower estuarine locations for the oyster beds rather than deeper open sea beds. The most likely scenario for Heybridge is that oysters were collected locally in the Blackwater estuary. In 1867 the upper reaches of the Blackwater owned by the Fish and Oyster Breeding Company were considered suitable for both breeding and fattening oysters but, at that time, shelly cultch had to be placed on the mud.
Comparisons of size and infestation characters with samples from other Roman sites indicate that the Elms Farm samples are typical of the east coast type of oyster (cf. south coast type of oyster) but are distinct from other Roman oysters found at North Shoebury and Colchester. This suggests that oysters from each of these three Essex sites came from different locations. In comparisons of size and infestation of Elms Farm oysters with modern oyster populations the results are equivocal.
Collection could have been by hand at low tide; the area in the upper Blackwater available for oysters in the late 19th century is known to have included acres of ebb-dry foreshore between the low water mark of neap and low water mark of spring tides. Fishing for oysters may have also taken place from boats using some kind of dredge, for which activity there is plentiful documentation in the 19th and 20th centuries but not earlier (Laver 1916). The oysters could then be ferried by boat and/or cart to a marketing point such as the supposed market area (Area H) of the Roman settlement, from which they could be transported far and wide.
Comparisons of both size and infestation yielded the interesting fact that 2nd-century oysters from context 15515 at Heybridge were virtually identical to oysters of the same date from context 1728 at Pudding Lane in London. The similarity is such that the two samples could have derived from the same oyster bed. This in turn suggests the possibility that oysters fished near Heybridge were traded on to London. It has already been suggested that the supposed relative paucity of oyster shells at Heybridge might be the result of selling on oysters to places such as Colchester and Chelmsford (Atkinson and Preston 1998, 108). This evidence suggests that oysters were traded further afield and introduces the idea that river and coastal transport may have been the logical way to get supplies to London.
Initially, perhaps, the live oysters would be carried just loose in sacks or baskets for movement from shore to market. However, greater distance and travel time would mean that the oysters had to be prevented from deteriorating or dying. While there are records of live oysters being packed in wooden tubs or bags tightly packed with snow to keep them cool as they travelled overland (Aldorovandi in Rydon 1968; Poli Testacea Utriusque Siciliae, quoted in Philpots 1890), it is also possible that they were transported in deep-welled boats around the coast.
Although Frere (1978, 337) suggests that oysters in Roman Britain were transported alive in tanks of water, in the same way that fish were kept alive, this would not have been an effective system. There is a high shell to meat ratio, i.e. a lot of waste, in a relatively low-cost commodity. Transport in tanks of water would be expensive and the numbers of oysters that could be carried this way would be limited. Additionally, it would have been difficult to maintain the quality of the salt water, which would easily stagnate and kill the oysters. Traditionally, live oysters have been packed tightly together, with the cupped left valve lowermost, in sacks, barrels or boxes so that the oysters have been prevented from opening their valves. This conserves the living animals in their own reservoirs of fluid. In cool conditions, well-packed oysters can survive at least ten days. Transport would be easier and costs would be far lower with oysters packed 'dry'. Transport and trade, both inland and coastal, would therefore have been highly likely.
At Elms Farm notches and cut marks were found on a few of the shells, which shows that at least some oysters were opened while alive and possibly eaten raw. Some of the shells had a brown iridescence on the internal surface, which is thought to be caused by burning, and other shells were blackened, both possible signs of cooking in the shell. If oysters are to be cooked, then roasting them in hot ashes or ovens causes the valves to open automatically as the animal is poached in its own liquor.
It can be assumed that after oysters have been transported some distance, whatever the mode of transport, by the time they reach their destination they will be in poor condition. Despite the Romans extolling the virtues of fresh oysters, there are many recipes for oysters and other shellfish cooked in rich and spicy sauces that would have masked any delicate natural flavour, and disguised many a bad one (e.g. Apicius translation Flower and Rosenbaum 1958; Edwards 1984, 82, 249, 257; Vehling 1977).
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Terms and Conditions | Legal Statements | Privacy Policy | Cookies Policy | Citing Internet Archaeology
Internet Archaeology content is preserved for the long term with the Archaeology Data Service (ROR). Help sustain and support open access publication by donating to our Open Access Archaeology Fund.