Cite this as: Murphy, P. 2015 Plant macrofossils from waterlogged contexts, in M. Atkinson and S.J. Preston Heybridge: A Late Iron Age and Roman Settlement, Excavations at Elms Farm 1993-5, Internet Archaeology 40. http://dx.doi.org/10.11141/ia.40.1.murphy
Thirty-eight samples from waterlogged fills of wells and a palaeochannel, dating from the 1st to 4th centuries AD, were examined. Low densities of cereal remains were present in most contexts, but were common only in the palaeochannel, where crop-processing waste was abundant. Other crops and utilised plants included dill, coriander, walnut, flax/linseed, possibly sainfoin, opium poppy and deadly nightshade; hazel, sloe, bramble and elder may also have been consumed. Macrofossils of dryland herbs, principally weeds and grassland species, were abundant. Trees and shrubs were sparsely represented. Despite the low-lying, poorly drained location of the site, close to an estuary, remains of wetland and aquatic plants and halophytes were rare. Assemblage composition was, overall, very consistent at all periods, and in different parts of the site. The results are compared with other sites in Essex.
The majority of cut features at this site were comparatively shallow, with well-drained fills of reworked gravel and brickearth. An extensive series of bulk samples was collected for retrieval, assessment and analysis of charred plant remains and other macrofossils. This report is concerned exclusively with plant macrofossils from the wet or waterlogged basal fills of deep pits, wells and a palaeochannel, ranging in date from c. 60-400+ AD.
Thirty-eight samples from the waterlogged fills were assessed. A small sub-sample (100-200g) was removed from each sample and disaggregated by manual agitation in hot water, before separating the organic fraction by wash-over, using a 0.5mm collecting mesh. The organic fraction was then scanned under a binocular microscope at magnifications of up to x50. In approximate chronological order the samples assessed were:
Well 13883 (Group 595, Contexts 18200, 18216, 18236, 18237, 18240). Area I, Period 3B, mid-1st to early 2nd century.
Well 9421 (Group 772, Contexts 9894, 9895). Area D, Period 3B, 2nd century.
Well 8188 (Group 788, Contexts 8167, 8214, 8195, 8196). Area E, Period 3, mid-2nd to early 3rd century.
Pit 9029 (Group 783, Contexts 8009, 9028, 9064). Area D, Period 3B, mid-2nd century.
Well 6280 (Group 531, Context 16074). Area H, Period 3B, mid- to late 2nd century.
Palaeochannel (Group 970, Contexts 12142, 12101, 12135). Area R, Period 4, 3rd century.
Well 14984 (Group 710, Contexts 14939, 20034). Area L, Period 4-5, early to mid-4th century.
Of these samples, seventeen produced macrofossil assemblages considered to be sufficiently large and well preserved for analysis (see Table 176, Table 177, Table 178, Table 179 and Table 180) (.xlsx). Samples selected for analysis were processed using the methods of Kenward et al. (1980).
Identifications were confirmed by comparison with modern reference material. Macrofossils of most taxa were counted, apart from Juncus seeds, whose abundance was assessed on a three-point scale. However, no counts were made of macrofossils from 12101, 20-30 and 30-40cm (Table 179) (.xlsx), for they appeared essentially identical to those from 12101, 10-20 and 40-50cm.
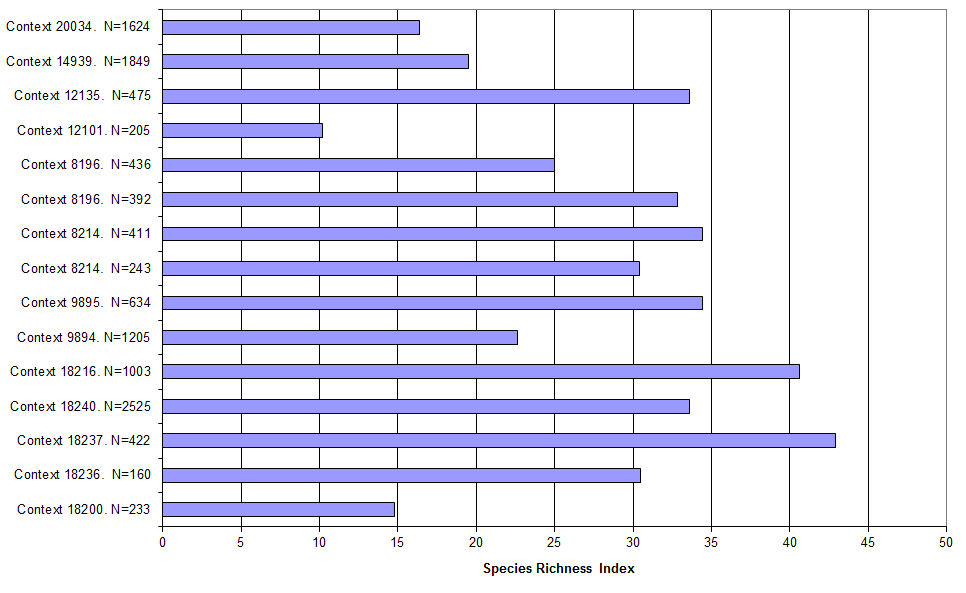
A Species Richness Index was calculated for each sample, as follows:
a/b x 100
Where, a = Number of taxa in sample; b = Number of taxa in whole assemblage (Wiltshire and Murphy 2004). At this site, b was taken as the total number of taxa from the site as a whole (Figure 687): 128 taxa in total.
It is well known that macrofossil assemblages from Roman and later contexts at urban and semi-urban sites are taphonomically complex, commonly consisting of inputs from more than one source. Assemblages composed of material from one source only are most uncommon. Interpretation of macrofossil assemblages from such sites is largely dependent upon understanding their taphonomy, and a simple Species Richness Index (SRI) provides an objective and quantified contribution towards this understanding (Figure 687). Assemblages which included a very wide range of taxa, and hence a high SRI are likely to have come from deposits that accumulated in situations where plant macrofossils from several sources were incorporated into them. By contrast, a low SRI can indicate a single source. However, it can also indicate poor preservation: for example, at the present site, a sample from the palaeochannel fill 12101 included only a restricted range of uncharred plant macrofossils (Table 179) (.xlsx), principally because it was an upper, partly de-watered, fill in which only durable macrofossils could be expected to be preserved: the SRI, 10.2%, was the lowest calculated for wet samples from this site.
In all other samples examined here, preservational factors are not thought to be relevant. In the well fill 18200 SRI was low (14.8%) but preservation good. The assemblage from this particular sample is thought to consist mainly of hay, with only a minor input from other sources (see below for further discussion). Samples from the early to mid-4th century Well 14984 (14939, 20034) also had low SRIs (19.5% and 16.4%) despite good preservation conditions. These two samples produced no residues from cultivated plants, aquatic taxa or coastal plants and very few macrofossils of scrub species: the assemblages from them were overwhelmingly dominated by weeds, mostly Urtica dioica (nettles): 90.4% and 91.9% respectively. This implies abandonment of the immediate area at this time, with dense development of weed vegetation; indeed no obvious sign of human activity at all.
For purposes of interpretation, the taxa identified have been grouped, partly in terms of the means by which they were preserved, partly on ecological grounds (Table 181 (.xlsx); Figure 688).
A very extensive series of bulk samples was collected for analysis of charred cereals and associated weeds, but these samples from waterlogged contexts also included charred material. In general, quantities were small. Many samples included a few charred caryopses and chaff fragments, but these are considered to be no more than the type of 'settlement noise' characteristic of Roman sites. However, fills of a palaeochannel in Area R, dating to the 3rd century included very high densities of charred cereal remains (Table 179) (.xlsx). This charred material consisted predominantly of glume bases and other chaff fragments of spelt wheat (Triticum spelta), with traces of emmer (Triticum dicoccum). Cereal grains were also present, together with fragments of plumules and radicles ('sprouts'). Occasional scraps of barley rachis internodes (Hordeum sp.) and wild or cultivated oat awn (Avena sp.) were also present.
Plainly crop processing waste accumulated, or was dumped, into this channel. The charred material is in a secondary context, so the precise activity represented is uncertain. However, at other low-lying Roman sites in Eastern England, where comparable assemblages have come from more secure contexts such as corn-dryers, they have been interpreted as relating to malting (e.g. Stebbing Green, Essex (Murphy 1999); Scole/Stuston By-Pass, Suffolk (Fryer and Murphy forthcoming). At these sites, cereal chaff was used as a fuel for parching malt: the resultant charred assemblages comprise a mixture of fuel residues and sprouted grains. Typically, these sites were in valley floor locations, adjacent to streams and rivers, where there was an ample water supply for steeping grain to initiate germination, and where discharge of waste water presented no problems.
The range of weed taxa represented by fruits or seeds is given in Table 181 (.xlsx). Even in the 3rd century palaeochannel fills, few taxa were identified. It is unclear how typical these might be of the arable weed flora in the area.
Several samples included a few uncharred glume bases of emmer, spelt and indeterminate wheat. These need not represent anything more than scraps of chaff blown about the site from areas where cereal processing was undertaken, or where cereal by-products were being used for flooring, litter, thatching or other purposes.
Taxa identified comprised Anethum graveolens (dill), Coriandrum sativum (coriander), Juglans regia (walnut), Linum cf. usitatissimum (flax/linseed), cf. Onobrychis viciifolia (sainfoin?), Papaver somniferum (opium poppy) and Atropa bella-donna (deadly nightshade) (Table 181) (.xlsx). In addition, some of the macrofossils of 'wild' trees and shrubs could represent food wastes, notably Corylus avellana (hazel), Prunus spinosa (sloe), Rubus section Glandulosus (bramble) and Sambucus nigra (elder). Macrofossils of cultivated plants other than cereals were, however, rare at Elms Farm, and no deposits rich in plant food residues were encountered.
Almost all of these taxa have previously been reported from Roman sites in Essex, notably from Colchester and the villa at Great Holts Farm, Boreham (Murphy 1984; 1992b; 1997b). The presence of Atropa bella-donna in Well 13883 (mid-1st to early 2nd century) calls for some comment. Deadly nightshade grows wild on well-drained chalk soils, and is most unlikely to have formed part of the local flora at Elms Farm. Seeds of this species have, however, been reported from a child's coffin, found preserved by waterlogging, on the line of the Scole/Dickleburgh By-Pass, Norfolk (Fryer and Murphy, forthcoming.). They were associated with leaves of box (Buxus sempervirens), and it seems highly likely that these were placed deposits, perhaps a wreath of box placed on the breast, with a strewing of deadly nightshade berries over the body. The possible symbolic attributes of these plants is obvious: box symbolising life, and deadly nightshade, death. Atropa therefore seems to have had some 'religious' associations, besides its pharmacological properties as a source of atropine.
The identification of Onobrychis viciifolia (sainfoin) was tentative, being based on a small fragment of tuberculate pod. It is a native plant (Godwin 1975, 180), growing principally on calcareous soils, but is also a fodder crop.
In addition to the scrub species mentioned above, samples included a few macrofossils of Betula sp. (birch), Salix sp. (willow/sallow) and Quercus sp. (oak). Seeds of Sambucus nigra (elder) were notably common in Well 9421 (2nd century), but the quantities observed could easily have come from a single bush. The macrofossils therefore provide little data on any possible phases of scrub development.
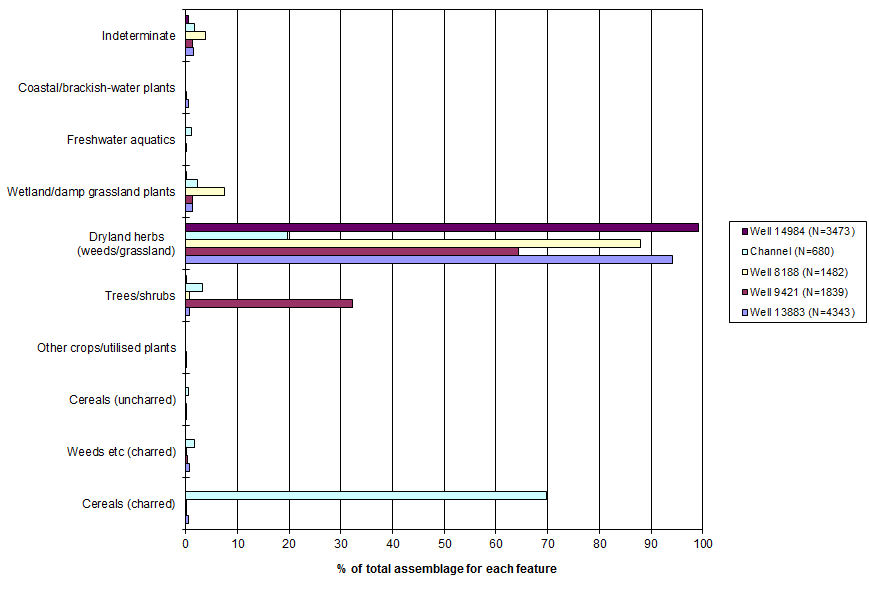
This 'ecological category' includes by far the largest number of taxa (Table 181 (.xlsx); Figure 688). Plainly, some taxa in this category are today exclusively grassland plants (e.g. Linum catharticum, Prunella vulgaris) and others occur exclusively in disturbed habitats (e.g. Papaver rhoeas, Thlaspi arvense). However, there is considerable overlap in the habitats of so-called 'weed' and 'grassland' plants, and this may have been still more marked in arable fields in the past, where tillage was relatively inefficient, permitting grassland species to persist (Hillman 1981). Equally, weed species commonly occur in areas of grassland disturbed and nutrient-enriched by grazing animals.
Greig (1988) has pointed out the very consistent composition of macrofossil assemblages from wells in the west and central Midlands, which he suggests indicates infilling by natural processes in areas of weedy abandoned land, while the surroundings were overgrown by scrub. Most of the assemblages from Elms Farm could be interpreted in this light. Alternatively, intentional back-filling with soil and other deposits from the immediate vicinity could be represented, for such deposits would have included abundant seeds and other remains of weed and scrub plants. Weed taxa occurring at notably high frequencies in at least some samples comprised Conium maculatum (hemlock), Chenopodium album (fat-hen), Rumex spp. (docks) and, ubiquitously, Urtica dioica (nettle). Segetal plants (arable weeds) such as Agrostemma githago (corn cockle) and Anthemis cotula (stinking mayweed) were not particularly common: it appears that local ruderal weed vegetation is principally represented, rather than residues from crop cleaning.
One sample, from context 18200 in the mid-1st to early 2nd century Well 13883, was rather different in character (Table 176) (.xlsx). It had a notably low Species Richness Index (14.8%), implying an origin mainly from one source. The dominant taxon was of small Poaceae (grass) caryopses (72.5% of total macrofossil count), which were associated with Leontodon sp. (hawkbit), larger Poaceae, Ranunculus acris/repens/bulbosus (buttercups), Eleocharis sp. (spike-rush), Filipendula ulmaria (meadowsweet), Juncus sp. (rushes), Rhinanthus sp. (yellow rattle) and Thalictrum flavum (meadow rue). 'Weed' species were only sparsely represented. Most of the plants represented in this sample fall into Greig's (1984) group of taxa characteristic of hay meadows, which are also likely to be represented in sub-fossil assemblages. It therefore seems probable that this sample either represented the immediate proximity of hay meadow or (more probably) incorporation of residues from hay in this well fill.
This group of plants was sparsely represented, compared to dryland herbs. Only Carex spp. (sedges), Eleocharis sp. (spike-rush) and Juncus spp. (rushes) were reasonably frequent, and then never abundant. Despite relatively high groundwater levels locally, the gravel surface appears to have been well drained, so that wetland and wet grassland vegetation did not develop in the vicinity. Even in the palaeochannel (Table 179) (.xlsx) wetland taxa were not common, implying that it was no longer active during infilling.
Obligate aquatic plants were conspicuously rare in samples from the wells at this site, and by no means common in the palaeochannel fills (Table 179) (.xlsx). Only three taxa were recorded: Alismataceae indet. (water plantain family), Lemna sp. (duckweed) and Ranunculus subgenus Batrachium (water crowfoot). Prior to back-filling, the wells may have been covered, and presumably any aquatic vegetation that did develop was removed. It appears either that there was little or no standing water in these features while they became infilled, or else that colonisation was not possible.
Despite the proximity of the estuary, coastal and brackish-water species were only sparsely represented. Only Triglochin maritima (sea arrow-grass) was at all frequent, notably in Well 13883 (mid-1st to early 2nd century), though rush seeds identified reasonably confidently (using criteria defined by Koerber-Grohne 1964) as Juncus gerardii (mud rush) occurred in Wells 13883, 9421, and 8188. Other taxa included Beta vulgaris (beet) and Daucus carota (wild carrot), both of which are found on embankments in the vicinity today, Oenanthe cf. lachenalii (parsley water dropwort?) and Scirpus maritimus (sea club rush).
It is plain that at no stage were conditions suitable for development of halophytic vegetation at the site: the very existence of the wells shows that groundwater was fresh throughout the Roman period. These macrofossils of coastal plants are therefore likely to have reached the site with plant material imported to the site for fodder, litter, thatching or other purposes.
Besides the fruits and seeds discussed above, most samples contained some other plant macrofossils, often not well preserved: charcoal, wood and twig fragments, buds and budscales, rosaceous thorns of Prunus/Crataegus and Rubus-type, mosses and, in Well 13883, pinnule and rachis fragments of Pteridium aquilinum (bracken). The rosaceous thorns were often associated with other macrofossils of Rubus and, occasionally, Prunus: they serve to demonstrate that thorny shrubs were growing locally. The bracken remains are likely to represent material imported to the site for use as litter and flooring materials.
Reference to Figure 688 illustrates the essential similarity of the assemblages from the wells, which date from the 1st to 4th centuries. In all cases, dryland herbs predominated, and most other groups were comparatively rare. In Well 9421, Sambucus seeds were common but, as noted above, this need not be significant. No local ecological changes are apparent. Charred cereal remains occurred in abundance only in the palaeochannel, at the northern edge of the main focus of settlement. It is reasonable to suppose that activities involving cereal processing were confined to such peripheral areas of the site.
Excavations at Slough House and Chigborough Farms, Heybridge, on the gravel terraces of the Blackwater, and in a very similar ecological situation to the site at Elms Farm, revealed a series of shallow wells/water-holes dating from the Bronze Age to the 7th century AD (Wiltshire and Murphy 1998). The essential difference, however, is that the Slough House and Chigborough Farm wells were not in areas of intensive settlement, but rather within field systems. They are thought to have been intended primarily for watering stock. Well 3887 at Chigborough Farm dated from the 1st century AD. As at Elms Farm, the macrofossil assemblages from this feature were dominated by dryland herbs: the most abundant taxa were Chenopodium album, Poaceae, Polygonum aviculare, Ranunculus spp., Rumex acetosella, Rumex sp., Spergula arvensis, Stellaria media-type and Urtica dioica. Thus, as at Elms Farm, grassland and weed vegetation predominated locally. However, in Well 3887, U. dioica was nowhere near so overwhelmingly abundant as at Elms Farm: annual weed taxa were more common. As at Elms Farm, trees, shrubs and plants of wet soils were sparsely represented. Freshwater aquatic species, particularly Lemna sp. and Ranunculus subg. Batrachium, were, however, extremely common. Overall, it appears that soils were less nutrient-enriched around Well 3887, as might be expected in a location away from intensive settlement. Furthermore, on abandonment, Well 3887 seems to have been left to infill gradually, providing an opportunity for aquatic vegetation to become established. By contrast, the wells at Elms Farm, located as they were within a settlement area, are more likely to have been rapidly back-filled when they went out of use.
Remains of crop plants were rare in Well 3887 (only a few wheat glume bases and a flax/linseed capsule fragment were identified). However, as noted above, they were by no means abundant in the wells at Elms Farm: there was little evidence from the site for disposal of domestic food wastes. This is in very marked contrast to the assemblages from a Late Roman well at the villa at Great Holts Farm, Boreham (Murphy 1997b), which included macrofossils from a wide range of fruit and nut crops. The Boreham assemblages appeared to represent waste flooring material (hay and straw) incorporating food waste. The complete absence of such dense deposits of domestic waste at Elms Farm is surprising, given the number of samples examined, and the location of the features sampled.
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Terms and Conditions | Legal Statements | Privacy Policy | Cookies Policy | Citing Internet Archaeology
Internet Archaeology content is preserved for the long term with the Archaeology Data Service (ROR). Help sustain and support open access publication by donating to our Open Access Archaeology Fund.