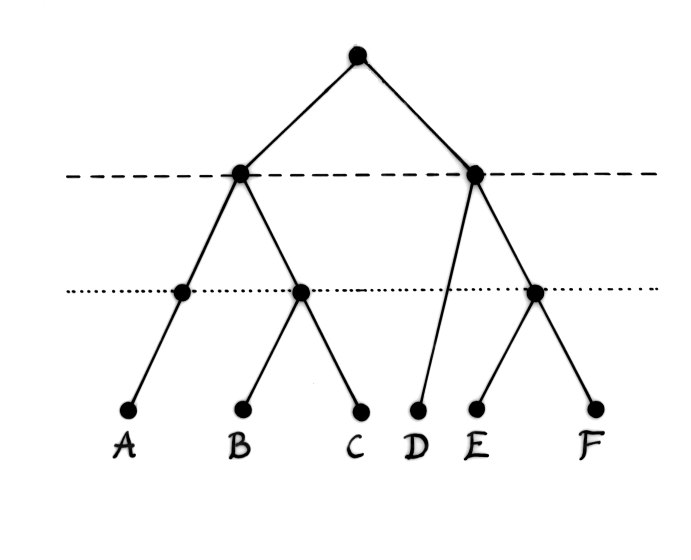
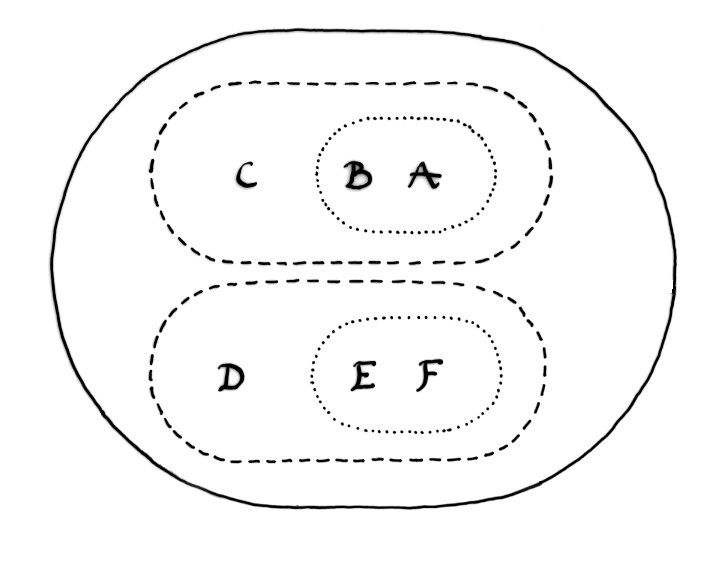
Figure 2b: The Venn diagram (b) represents the classification obtained by analysing the hierarchic structure.
The palaeoanthropological evidence, reviewed in detail elsewhere (Winder and Winder 2014) is not consistent with the model of evolution as a process of divergence between reproductively closed populations. Indeed, if a clear hierarchy could have been characterised, the problems of human origins would have been solved long ago. Rather, the fossil evidence suggests a patchily distributed, genetically open population of great apes organised into close-knit pro-social units.
Modern primates sometimes have complicated sex lives. Hybrids between species and, on occasions, between genera have been observed, both in captivity and within wild populations. It is easy to see how these traits, complemented with matings between close kin, would facilitate survival among tiny populations driven through demographic bottlenecks. These are situations where conventional mating strategies and barriers to hybridisation would lead to extinction. In order to survive, individuals must mate with close kin or, on occasion, with members of another species. The result would be a braided stream or heterarchy of crossing and re-crossing lineages in which a mosaic of morphological and physiological traits were manifest (see Figures 2 and 3 & Winder and Winder 2014). Darwin's solution to the Goldilocks problem would not work in these circumstances because upwellings of hidden traits would destabilise established attractors.
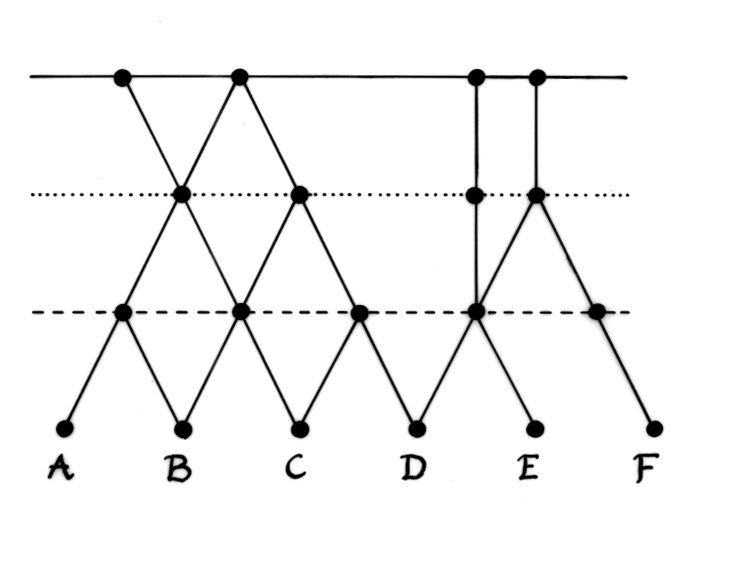
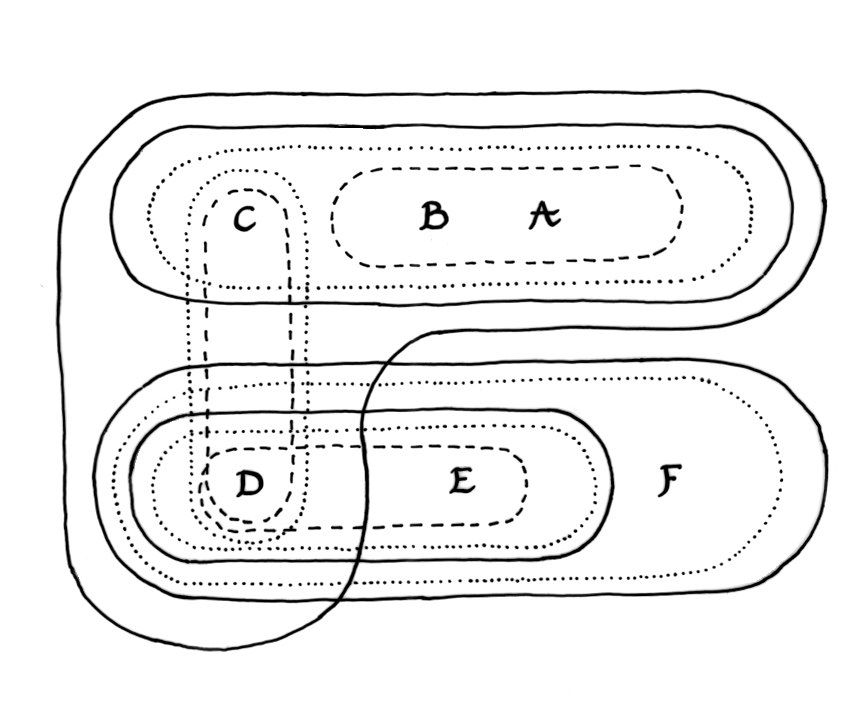
The effect of these upwellings would be to create a meta-selection pressure that favoured plesionic systems able to acquire coping strategies. Strong dominance/recessive relations and modifier genes that could prevent the expression of deleterious traits are common biomolecular coping strategies. However, behavioural flexibility, social learning and the ability to innovate may also have been significant. There is a growing body of archaeological and primatological evidence (Hublin 2009; Spikins et al. 2010) consistent with the hypothesis that populations which were a little smarter and more compassionate would be able to mitigate the effects of natural selection faster and discover new ways of becoming synergetically fit again.
The last 6-7 million years have seen more positive selection in the chimpanzee line than our own (Bakewell et al. 2007), a result consistent with the hypothesis that chimpanzee lineages did not acquire the flexible, co-operative strategies needed to hide deleterious genes from selective winnowing. Our ancestral lineages seem to have formed a braided stream of crossing and re-crossing flows. These flows would have converged and deepened into a small number of recognisable species, many of them capable of flexible social learning, pro-social, empathic, and compassionate.