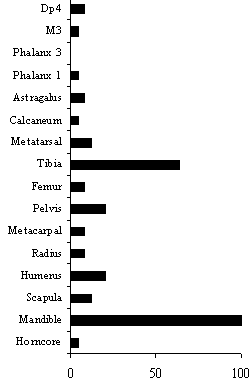
Fig.66: Welton Road, Brough: caprovid MNI by anatomical element (see Table 40a for data)
The relative abundance of the different skeletal parts recorded in an archaeological assemblage can be used to interpret a range of human activities relating to the processing and disposal of the carcasses (Dobney et al. 1996a, 23).
Table 40a-b and Figs.66-7 illustrate the relative abundance of caprovids and cattle skeletal elements. The sample size remains very small, even after the data from Phases 1 and 2 have been combined. To minimise the problems created by the small sample size the simplest method of calculating the relative abundance of body parts has been used. The MNI of all skeletal elements is expressed as a proportion of the most frequent element (shown as 100%). For these tables and figures MNI values have been calculated by assuming there are equal numbers of left and right elements.
| Element | No. of fragments | MNI | % |
| Horncore | 1 | 1 | 4 |
| Mandible | 50 | 25 | 100 |
| Scapula | 6 | 3 | 12 |
| Humerus | 10 | 5 | 20 |
| Radius | 4 | 2 | 8 |
| Metacarpal | 4 | 2 | 8 |
| Pelvis | 10 | 5 | 20 |
| Femur | 3 | 2 | 8 |
| Tibia | 31 | 16 | 64 |
| Metatarsal | 6 | 3 | 12 |
| Calcaneum | 2 | 1 | 4 |
| Astragalus | 4 | 2 | 8 |
| Phalanx 1 | 5 | 1 | 4 |
| Phalanx 3 | 0 | 0 | 0 |
| M3 | 2 | 1 | 4 |
| Dp4 | 3 | 2 | 8 |
| Element | No. of fragments | MNI | % |
| Horncore | 2 | 1 | 9 |
| Mandible | 22 | 11 | 100 |
| Scapula | 4 | 2 | 18 |
| Humerus | 9 | 5 | 45 |
| Radius | 4 | 2 | 18 |
| Metacarpal | 16 | 8 | 73 |
| Pelvis | 6 | 3 | 27 |
| Femur | 1 | 1 | 9 |
| Tibia | 6 | 3 | 27 |
| Metatarsal | 15 | 8 | 73 |
| Calcaneum | 12 | 6 | 55 |
| Astragalus | 15 | 8 | 73 |
| Phalanx 1 | 78 | 10 | 91 |
| Phalanx 3 | 18 | 2 | 18 |
| M3 | 4 | 2 | 18 |
| Dp4 | 2 | 1 | 9 |
Fig.66: Welton Road, Brough: caprovid MNI by anatomical element (see Table
40a for data)
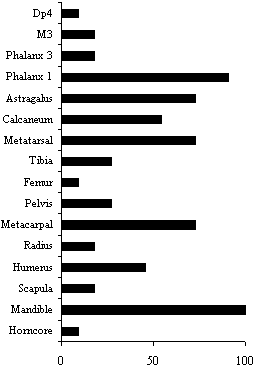
Fig.67: Welton Road, Brough: cattle MNI by anatomical element (see Table
40b for data)
It is immediately clear that the assemblage is biased against the major meat-bearing bones and in favour of the distal limb bones and mandibles. There are two possible explanations; firstly that this reflects the greater durability of certain dense bones, and secondly that this reflects intensive exploitation of the carcass and the disposal of selected body parts. In reality, it is probably a combination of both.
The pattern of butchery has many of the distinctive features associated with the organised, large-scale processing of cattle carcasses in the Roman period (Hamshaw-Thomas and Bermingham 1993; Dobney et al. 1996a; Maltby 1989; Smith 1996, 20). Evidence from the bones suggested that much of the processing of cattle carcasses was achieved by the use of a heavy chopper. Knife marks were only recorded on the phalanges, presumably resulting from their removal from the hide. A number of the epiphyses and the tarsal bones bear witness to the systematic reduction of the carcass into small pieces for further processing, either for a broth (Mensch 1976) or, more likely, marrow extraction. There was clear evidence of this process in the form of a number of longitudinally split long bones. The many transversely broken metapodial fragments were also probably the result of marrow extraction. These processes will have reduced the representation of the meat-bearing bones in Figure 67.
Interestingly, several mandible fragments were split or chopped through the diastema. Four diastema fragments from context 1045 were split and burnt. This feature has been interpreted as a result of the extraction of liquid marrowfat for lamp oil or as a base for cosmetics (Dobney et al. 1996a, 26). No cattle scapulae with perforated blades were recorded, although examples of the removal of the spina was noted.
The almost total lack of horncore fragments suggests that the horncores were perhaps removed and processed at some other location. Negative evidence is, however, dubious and a number of taphonomic reasons could account for the lack of horncores. One horncore fragment showed signs of being worked. The small curved rectangular fragment (approximately 4 x 2.5cm) had a chopped and sawn edge, suggesting the unsuccessful production of a blank.
Despite the small sample size, it is possible to see that processing of the carcasses was well organised and followed a similar pattern to that deduced for contemporaneous sites. Primary butchery appears to have taken place on or near to the site. The dressed carcasses were then further reduced, in some cases very intensively. There is little evidence for domestic or kitchen refuse which suggests that this type of waste may have been dumped elsewhere on the site. Alternatively, meat may have been exported to other locations, with the remains of the main domesticates from Welton Road, Brough, reflecting a producer rather than a consumer economy.
The caprovid sample is less easy to interpret. A similar pattern to that for the cattle could be envisaged if one takes into account the possibility that the smaller distal limb bones were not recovered. However, the samples that were processed did not contain large quantities of these elements. It is safer, therefore, to view the skeletal element representation as a reflection of the small sample size and the variable durability and archaeological visibility of the different body parts.
The pattern of cut and chop marks on the caprovid bones is unremarkable. As with the cattle, no scapulae had perforated blades, although there was again evidence for the removal of the spina in some cases.
As previously mentioned, four of the horse bones bore signs of butchery. The position of the cut marks was consistent with the dismembering, skinning and filleting of the carcasses. Whether horses were actually eaten cannot easily be established.
Butchery marks were also recorded on the lingual surface of a pig mandible, presumably the result of the extraction of the tongue.
© Internet Archaeology
URL: http://intarch.ac.uk/journal/issue9/brough/eaurep4.html
Last updated: Tue Nov 28 2000