Seven 14C dates on procellariid bone from sinkhole sites
1710-1 and 9659-1 (Table 5) provide the data needed to construct
an absolute chronology for the stratigraphic sequences of Kalaeloa sinkholes (14C age determinations are listed fully here because, to the best of
our knowledge, all previous listings are either incomplete or contain errors;
the information in Table 5 is from records kept at Beta Analytic dating
laboratory).
Davis designed the dating project carefully, selecting bone samples for dating
from each of three depositional units present at the two sites. Because of
this, the dating calibration can integrate relative stratigraphic information
by use of a Bayesian statistical framework (Buck et al. 1996; 1992; 1991). In
this framework, information on relative ages of dated events is used to
constrain the calibrated ages of dated samples; the calibrated age of a sample
will always be younger than the calibrated age of a sample recovered from a
stratigraphically older deposit, regardless of the relative 14C ages
of the two samples. Thus, samples that yield inverted 14C ages
are restored to their correct relative ages, as this relationship is defined
stratigraphically. Typically, addition of stratigraphic information to the
calibration procedure improves the ability to interpret the results archaeologically.
In addition, adoption of a Bayesian framework provides a way to obtain age
estimates for events that were not directly dated, which is useful in this
case because it is possible to estimate ages of depositional unit boundaries.
Under the assumption that changes in depositional modes were
penecontemporaneous across the region, age estimates for depositional unit
boundaries derived from sites 1710-1 and 9659-1 can be extrapolated to
depositional sequences of sinkhole sites that were not dated. A primary
objective of the Bayesian 14C calibration reported here is to estimate calendar
ages of transitions from one depositional unit to the next.
Lab. no.
Site
Depth (cm)
Dep. unit
CRA
δ13C
Event
Beta-11192
9659-1
10-20
transported
920±100
-17.6
θ1
Beta-11193
9659-1
20-30
collapse
1130±100
-17.5
θ2
Beta-11194
9659-1
30-40
collapse
1370±100
-19.0
θ3
Beta-11188
1710-1
16-26
collapse
1090±100
-23.2
θ4
Beta-11189
1710-1
26-36
collapse
1260±100
-13.8
θ5
Beta-11190
1710-1
36-46
collapse
1730±100
-19.8
θ6
Beta-11191
1710-1
56-65
basal
2320±100
-24.7
θ7
Table 5: 14C dates on procellariid bone
The analysis requires three assumptions: 1) sediment
deposition in sinkholes is continuous - there are no hiatuses between or within
depositional units; 2) there is no significant hiatus between the death of the
bird whose bone was dated and deposition of the bone in the sinkhole; and 3)
post-depositional movement of dated bones through the stratigraphic column was
not sufficient to change their positions relative to depositional unit
boundaries. We believe that these assumptions are reasonable at this stage of
analysis, but do not believe that they are universally valid. In our view,
further study of the dynamics of sediment deposition in sinkholes is clearly
warranted.
Given the stratigraphic and 14C information, and assumptions
listed above, it is possible to formulate a model of the relationships among
depositional units and unknown calendar ages of events represented by seven
14C dates. Following standard practice, we indicate
the lower boundary of depositional unit i (i = I, II, III) as βi and the upper boundary as αi. Let θj denote the
unknown calendar date BP of event j (j = 1..7). Then archaeological and 14C information from the two Kalaeloa sinkholes can be expressed in the form of the following inequalities.
This model was implemented using the OxCal software package
(Ramsey 1995). Seven 14C determinations associated with the θi (Table 5) were calibrated with a marine curve (Stuiver and
Braziunas 1993) using a δr value
of 110±80 established for ocean waters
surrounding the Hawai'ian Islands (Dye 1994b).
Event
Davis
Christensen
Dye & Tuggle 95.4% h.p.d.
θ1
AD 1265-1490
AD 1030
AD 1420-1880
θ2
AD 1215-1410
AD 820
AD 1150-1510
θ3
AD 1030-1325
AD 580
AD 890-1340
θ4
AD 1255-1415
AD 860
AD 1200-1540
θ5
AD 1055-1350
AD 690
AD 1010-1410
θ6
AD 790-1215
AD 200
AD 580-1060
θ7
AD 445-855
370 BC
200 BC-AD 450
Table 6: Estimated ages of dated events. Sources: Davis (1990); Christensen (1995)
Estimates of the calendar ages of the dated events are
listed in Table 6 as 2 σ highest posterior density
regions, along with calendar ages reported by Davis (1990) and
Christensen (1995). The estimates yielded by Bayesian analysis are younger by
200-700 years than age estimates reported by Christensen (1995), as expected.
They are, however, very close to the results yielded by Davis' calibration
procedure, with two important exceptions. The two exceptions are θ7, at the early
end of the sequence, and θ1 at the
late end of the sequence. The Bayesian estimate for the age of θ7 is 400-600 years
earlier than Davis' estimate, and the estimate for θ1 is 200-400 years later. These differences have
the effect of doubling the estimated duration of the interval between the
earliest and latest events in the sequence, transforming the 400-1,000 year
sequence posited by Davis, to one that spans 1,000-2,000 years.
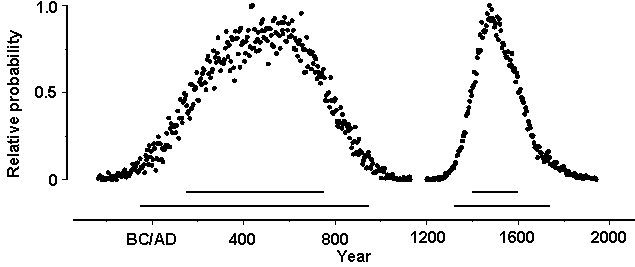
Figure 3: Estimated ages of depositional unit boundaries. Left, boundary of basal diagenetic and structural collapse deposits; right, boundary of structural collapse and transported sediment deposits. Solid lines above the x-axis indicate 67%
and 95.4% highest posterior density regions
The 95.4% highest posterior density region yielded by
Bayesian calibration for the estimated age of the boundary of basal diagenetic
and structural collapse deposits is 50 BC-AD 950 (Fig. 3), an interval that spans current estimates of the date
of initial Polynesian colonisation of the islands. The date of colonisation
has become a point of contention, over which roughly two schools of thought
have formed. There is an argument for 'early' colonisation dating to the AD
100-400 range (e.g. Kirch 1985; Hunt and Holsen 1991) and an argument for a
'late' colonisation, as late as AD 600-1000 or even AD 800-900
(e.g. Spriggs and Anderson 1993; Athens et al. 1997). Events in the basal
diagenetic deposit, exemplified by θ7, are likely to have occurred either very early in
the Polynesian era, or before Polynesian colonisation of the islands. Although
the early colonisation estimate is coeval with the boundary of basal
diagenetic and structural collapse deposits, it is unlikely that Polynesians
would have settled or farmed this marginal region soon after colonization
(Tuggle 1997) and it is safe to say that events in basal diagenetic deposits
pre-date significant Polynesian activities at Kalaeloa. It is not possible
with the data at hand to estimate with confidence when basal diagenetic
sediments were first deposited. This event pre-dates event θ7, however, and a
reasonable inference is that deposition of basal diagenetic deposits began
more than 2,000 years ago.
The 95.4% highest posterior density region for the estimated
age of the boundary of structural collapse and transported sediment deposits
is AD 1320-1740, an interval that encompasses
the last four centuries of the pre-Contact era (Fig. 3). The structural collapse deposits represent at least
the first half of the Polynesian era, but given uncertainties in the date of
Polynesian colonisation and in the age estimate of the depositional unit
boundary, these deposits might encompass nearly the whole of the era. Events
in transported sediment deposits either took place late in the pre-Contact
era, or in the period after Contact. The bone dated for event θ1, whose estimated
age falls late in the Polynesian period, was collected from the bottom half of
the transported sediment deposit at site 9659-1 and is consistent with this
assessment.