5.2 Changes in populations of native, extinct taxa
The primary datum in the argument for
extinction by Polynesians of land snails at Kalaeloa is a decline over time
in relative proportion of 18 native taxa in excavated archaeological and
palaeontological collections. This pattern was first noted by Christensen and
Kirch (1986) at site 9574, where native, extinct taxa are absent in leaf
litter on the surface of the sinkhole. Subsequent excavations and analyses
failed to yield another site where the decline of native, extinct snails was
complete. At several sites this decline is pronounced (e.g. 9647-2, 9661-2),
but at others the decline is more moderate (e.g. 1710-1, 9659-1, 9670-P1). At
most sites the relative proportion of native, extinct taxa never declines to
nil. Native, extinct taxa are absent only in the cultural layer (II) at site
2701-1 and the leaf litter of sinkhole site 9574.
Given the interpretative difficulties inherent in use of
relative proportions, we analysed change over time in diversity of land snail
collections, using measures of richness and evenness to characterise
diversity. Richness is a straightforward measure that can take on integer
values between 0 and the maximum number of taxa in the collection. Evenness is
more difficult to quantify and several different measures have been proposed
(Bobrowsky and Ball 1989). The measure used here, called H/Hmax by
Kintigh (Kintigh 1989, 29), can take on real values between 0.0 and a maximum
that depends on the richness of the collection, but usually is between 2.0 and
3.0. In an extinction event, both richness and evenness decline to 0.
The richness and evenness measures on Kalaeloa snail
collections estimate diversity of land snail faunas that existed within
catchments of sites where they were deposited. Like richness measures,
evenness measures are sensitive to sample size, and both measures can yield
poor estimates of faunal diversity if sample sizes are small (Grayson 1984, 158
ff.). The Monte Carlo technique used to estimate expected richness, discussed
above, can also estimate expected evenness (Kintigh 1989) and estimates of
expected richness and evenness were calculated for each of the land snail
collections, using the site collection as the parent population from which
samples were drawn. Use of site collections, rather than the Kalaeloa
collection, as the parent population, helps to control for differences in
diversity of land snail faunas among different habitats.
Figures 4-9 compare relative proportions of native, extinct taxa
over time with their expected richness and evenness at six sinkhole sites.
These include two sinkhole sites analysed by Christensen and Kirch (1986) and
four of eight sinkholes excavated by Davis (1990) for which land snails were
identified (Christensen 1995); sediments of these four sinkholes were analyzed
by Allen (1995) and it is possible to assign estimated ages to their
depositional unit boundaries. Error bars for relative proportions are based on
one standard deviation counting errors, calculated as square root of sample
size. Richness and evenness are expressed on the figures as the difference
between observed and expected values generated by the Monte Carlo method.
Error bars for richness and evenness represent 80% confidence intervals. It is
rare at Kalaeloa to find a collection that is as diverse as expected by the
Monte Carlo method, and most of the plotted differences between observed and
expected diversity measures are negative. This is because most sinkhole sites
show evidence for change over time in the land snail fauna, including episodes
of colonisation and extirpation of taxa, which affect richness, and changes in
relative proportions of taxa, which affect evenness. Because any single
collection contains material from only a portion of the time that materials
were deposited in the sinkhole, use of the site collection as the parent
population in the Monte Carlo method often overestimates diversity of
individual collections.
The distributions of introduced taxa L. gracilis and G. servilis
are illustrated in the bottom panel of figures because they were used
by Christensen and Kirch (1986) as index sub-fossils, as discussed above. They
are plotted on a logarithmic scale as MNI + 1, to compensate for wide
variation in counts from one collection to the next, many of which include nil
values.
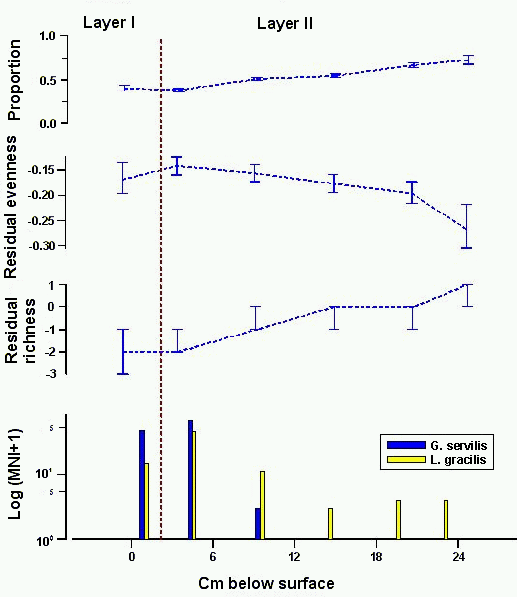
Sinkhole site 9574 is the type site for a decline in
native, extinct land snails at Kalaeloa. Christensen and Kirch (1986) divided
the 85cm deep stratigraphic column into three layers and collected ten
samples for land snail identification. Layer I, a thin, black, relatively
loose A0 and A1 horizon yielded a single sediment sample for land
snail identification; Layer II, a 30cm deep, brown, silt and sand deposit
with angular fragments of limestone yielded three samples; and Layer III, a 50cm deep, reddish yellow deposit of limestone breccia in a matrix of silt and
sand yielded five samples. A single sample collected from leaf litter above
Layer I represents the modern land snail fauna of the sinkhole.
Christensen and Kirch (1986) argue for a period of increased
change and decline in relative proportion of native, extinct taxa beginning in
Layer II and continuing through to their extirpation. This argument is based
on the following data (Fig. 4, top). Native, extinct taxa make up 71±3% of the land snail collection in the bottom sample
from Layer III at the base of the stratigraphic column and rise to a maximum
of 85±6% in the top two samples from this layer.
This high level is maintained through the bottom two samples in Layer II, but
it declines significantly in the top sample to 67±2%. This proportion is not significantly different
from the proportion at the base of the stratigraphic column and the relative
proportion of native, extinct taxa in Layers II and III can be characterised
as fluctuating slightly between 65% and 91% of the total collection. The
relative proportion of native, extinct taxa does decline significantly in
Layer I, where they make up only 20±1% of the
collection. There are no native, extinct taxa in the leaf litter (native, extinct taxa were found in leaf litter at sites 9647-2 (Fig. 8) and 9661-2
(Fig. 9)).
Figure 4: Native, extinct land snail taxa at sinkhole site 9574. Top, relative proportion with 1 errors; second from top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale) Analyse the data using XLisp-Stat
The diversity measures for native, extinct taxa at site 9574
show no sign of a significant decline through three layers of the
stratigraphic column, and it is not until the leaf litter, when both measures
decline to 0, that diversity measures indicate any stress in the native,
extinct land snail fauna.
It is not possible to correlate stratigraphic layers with
Allen's depositional units and there are no 14C dates from the site, so
absolute dates cannot be assigned to changes in land snail collections.
Christensen and Kirch (1986) believed that Layer II was deposited, at least in
part, during the Polynesian period, based on occurrence of small numbers of
L. gracilis shells in the top two samples from
the layer. This inference is by no means secure, however, as discussed below.
The only layer that is securely dated is Layer I, whose rich collection of
Polynesian and historically introduced land snail shells indicate that it was
laid down after Polynesian colonisation and into the post-Contact period.
Christensen and Kirch (1986) correlated the onset of decline
in relative proportion of native, extinct taxa with Polynesian influence. This
inference was based on the stratigraphic distribution of various animals
introduced by Polynesians, including the land snail L.
gracilis. As the bottom panel in Figure 4
shows, however, correlation of the decline with introduction of L. gracilis is not
particularly clear. L. gracilis, is first present
in Layer II, but it occurs as five or fewer individuals in two Layer II
collections in which it appears, and is not well-represented until Layer I,
when the historically introduced taxon G. servilis
is also first present in large numbers. Given pervasive mixing of small
numbers of land snails down through the stratigraphic column at sites
excavated subsequently (Christensen 1995), the presence of a few shells of L. gracilis in Layer II does not necessarily indicate
this snail's colonisation of the sinkhole at the time Layer II sediments were
deposited. The distribution of G. servilis down
to the base of the stratigraphic column is apparently not the result of mixing, and
was explained as contamination of sampled sediments by living snails that
colonised the excavated portion of the sinkhole before land snail samples were
collected (Christensen and Kirch 1986, 63). (Curiously, L. gracilis is absent in the leaf litter along with native, extinct taxa.
The other introduced taxon, G. servilis, is the dominant component of the leaf litter
collection.)
The history of the native, extinct land snail fauna that
emerges from this analysis is very different from the history posited by
Christensen and Kirch (1986) based on a consideration of relative proportion.
The population of native, extinct land snails at site 9574 is stable until the
loose sediments of Layer I were deposited, at which time the relative
proportion of native, extinct taxa falls to levels not experienced previously.
This is stratigraphically coeval with colonisation of the sinkhole by the
historically introduced G. servilis, and possibly by the Polynesian introduction
L. gracilis, as well. The diversity of native,
extinct land snails shows no stress in the population at this time, but
declines suddenly after Layer I was deposited. This indicates that a drastic
change leading to extirpation of native, extinct taxa at the site belongs
either to the late pre-Contact period, or more likely to the post-Contact
period. There are no data here that indicate changes in the native, extinct
land snail population were due to Polynesian settlement, rather than to major
anthropogenic environmental changes to the region documented historically.
5.2.2: Site 9670-P1
Sinkhole site 9670-P1 was also analysed by Christensen and
Kirch (1986). The deposits at site 9670-P1 are much shallower than at site
9574, reaching a maximum depth of 27cm. The stratigraphic sequence at the
site is divided into three layers. One sediment sample for land snail
identification was collected from Layer I and five samples were collected from
Layer II. No samples were taken from Layer III.
Figure 5: Native, extinct land snail taxa at sinkhole site 9670-P1. Top, relative proportion with 1 errors; second from
top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale) Analyse the data using XLisp-Stat
Christensen and Kirch (1986) hypothesized, on the basis of
the distribution of L. gracilis to the base of the stratigraphic column, that
the stratigraphic sequence correlated with the upper portion of the sequence
at site 9574 (Christensen and Kirch 1986, 72). The land snail sequence at site
9670-P1 differs from the upper portion of the sequence at site 9574, however,
in that the relative proportion of native, extinct taxa declines moderately up
through the stratigraphic column and native, extinct taxa never disappear from
collections. Instead, they represent 40±3% of the
total land snail collection in Layer I. There is a slight, but significant,
decline in richness between the sample from the base of the stratigraphic
column and two samples from the top of the column, but there is no significant
change in evenness.
As was the case with site 9574, it is not possible to
correlate the layers at site 9670-P1 with the depositional units of
Allen (1995). The distribution of introduced taxa L.
gracilis and G. servilis through the
stratigraphic column is consistent with the relative timing of their
introductions, and supports the inference that Layer II was laid down from the
Polynesian period through to the early post-Contact era, although the small
number of L. gracilis shells in collections from
the bottom half of Layer II suggest some interpretative caution (pace Christensen and Kirch 1986, 64). A plausible
argument could be made for deposition of the bottom half of Layer II during
pre-Polynesian times, with subsequent downward mixing of a few L. gracilis shells. Lacking 14C dates
it is not possible to discriminate between these alternative hypotheses.
It is difficult to see in these data any effect on native,
extinct taxa that can be attributed to Polynesian influence. Christensen and
Kirch (1986) chose to focus their analysis on the decline in relative
proportion of just two native, extinct taxa (O. uberta
and E. kalaeloana) and increases in three
native, extant taxa (Lamellidea spp., Tornatellides spp., and S. caduca), citing these trends as supporting evidence for the decline in
native, extinct taxa documented at site 9574. The implication of this choice
is that differences in the depositional histories of land snails at sites 9574
and 9670-P1 reflect a deviation at site 9670-P1 from a widespread pattern,
rather than simply an expression of general variability in land snail faunas
among Kalaeloa sinkholes.
5.2.3:Site 9659-1
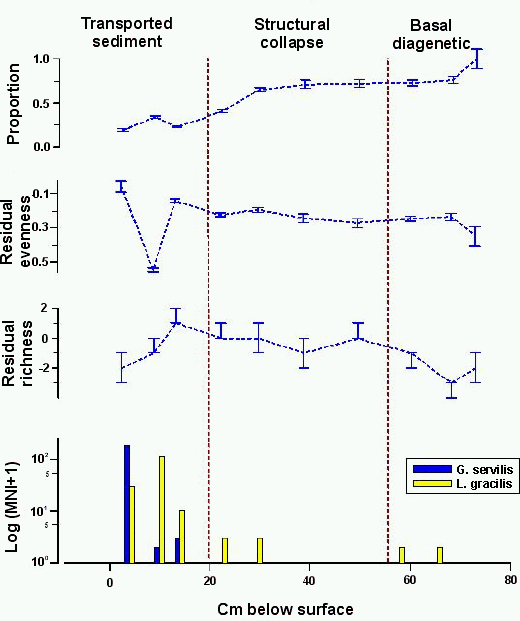
Site 9659-1 was excavated by Davis (1990). His analysis
indicated that all of the sinkholes he excavated at Kalaeloa 'yielded similar
patterns of change in the snail assemblage' (Davis 1990, 197) and that this
change was that described by Christensen and Kirch (1986) at the land snail
'type site', sinkhole site 9574. Davis (1990) illustrated the pattern of
change with the land snail sequence from an excavation under the overhang at
sinkhole site 9659-1, which he believed showed the pattern most clearly. The
~50cm thick sediment deposit was excavated in
seven arbitrary 5cm and 10cm levels that were later assigned to four layers
(Davis 1990, 182). Allen (1995) recognised three depositional units in the four
layers. A total of ten sediment samples (Christensen 1995, 245) was collected
for land snail identification. (The bar graphs in Davis (1995, 199) and Davis (1990, 438) indicate 15 and 13
samples, respectively, but there is no published record of samples identified in
addition to the ten identified by Christensen (1995).) The ~21cm thick
gravelly loam of the transported sediment deposit (Layers I and II)
yielded three sediment samples for snail identification; four samples were
taken from the 15-20cm of very gravelly, coarse sandy loam of the structural
collapse deposit (Layer III); and three samples were collected from the 5-10cm of gravelly, very coarse sandy loam of the basal diagenetic deposit (Layer
IV and Layer V from Square 4 in the exposed portion of the sinkhole).
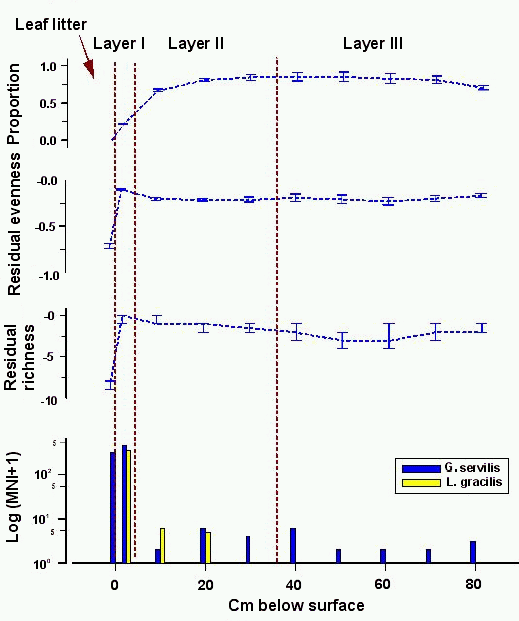
There are two periods of marked decline in relative
proportion of native, extinct taxa in the collections (Fig. 6), although the sequence itself is characterised by a
first-order decrease in relative proportion over time. The first period of
decline occurs at the base of the stratigraphic column where the proportion
drops from 100-11% to 76±4%. The second begins at
the top of the structural collapse deposit, where the relative proportion is
65±3%, and continues through to the base of the
transported sediment deposit where relative proportion drops to 22±1%. Native, extinct taxa are well-represented at the
top of the stratigraphic column, making up 19±2%
of the collection there. (Davis describes this collection as consisting 'entirely of native survivors and
more recently introduced snails' (Davis 1990, 198).) Use of the term 'survivors' is
potentially confusing here, as it seems to indicate that only native, extant taxa
remain. In fact, both native, extinct taxa and native, extant taxa are present in the
collection.
In contrast to the moderate first-order decline in relative
proportion of native, extinct taxa, richness and evenness measures indicate an
increase in diversity through to the base of the transported sediment deposit.
Richness declines in the transported sediment deposit and evenness dips deeply
in the middle of the deposit due to a sudden peak in N. newcombi shells, which make up 25.5% of the collection
(Christensen 1995, 246). These data indicate that there was some stress in the
population of native, extinct taxa by the middle of the transported sediment
deposit.
This sinkhole was dated by Davis and age estimates can be
assigned directly to depositional unit boundaries (Section 5.1). The distributions of introduced
taxa appear to substantiate the chronology derived from 14C dates.
G. servilis is found only in the transported
sediment deposit, and in significant quantities only in the top-most
collection, so the lower boundary of the unit appears to be late pre-Contact
in age. L. gracilis also occurs most frequently
in the transported sediment deposit, but is found in scant numbers in the
structural collapse and basal diagenetic deposits as well. Presence of this
taxon in deposits laid down in the era before Polynesian colonisation, and its
discontinuous distribution through the stratigraphic column (Christensen (1995, 245) reports non-apical fragments of L. gracilis from the
lower two samples in the structural collapse deposit) suggest that
the few shells in the basal diagenetic deposit are intrusive. Although the
four MNI of L. gracilis at the top of the
structural collapse deposit might indicate when this snail colonised sinkhole
9659-1, it is also possible that these few shells, like the few shells in the
basal diagenetic deposit, are intrusive. If this is the case, then L. gracilis was first deposited in transported
sediments, either late in the pre-Contact era or early in the post-Contact
period.
Figure 6: Native, extinct land snail taxa at sinkhole site 9659-1. Top, relative proportion with 1 errors; second from top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale)
The timing of changes in land snail collections at site
9659-1 casts doubt on the proposition that Polynesian activities were
responsible for the decline of native, extinct taxa. First, the relative
proportion of these taxa declines over the entire stratigraphic sequence.
Onset of this first-order decline is likely to pre-date the fifth century
AD (Fig. 3).
Second, diversity of the native, extinct land snail population does not
decline until after deposition of the transported sediments began. This
depositional unit began either very late in the pre-Contact era or in the
post-Contact period (Fig. 3). Finally,
regardless of when the population of native, extinct taxa began to decline,
either late in the pre-Contact era or in the post-Contact period, the cause of
decline was insufficient to extirpate them at the site. At the top of the
stratigraphic column, the population of native, extinct taxa is as diverse as
it was in collections from both Polynesian and pre-Polynesian periods.
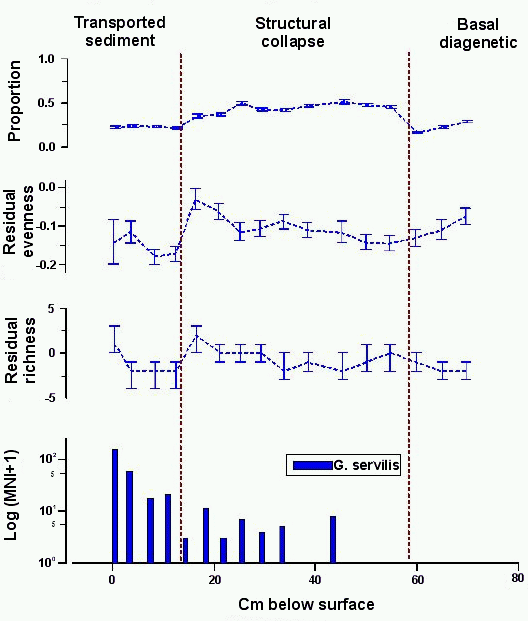
5.2.4 Site 1710-1
Sinkhole site 1710-1 was excavated in four layers to a
maximum depth of 76cm, comparable to the deep deposit in site 9574
(Davis 1995). Land snails were identified in sixteen sediment samples from the
site. The transported sediment deposit (Layers I and II), a dark, organic
loam, yielded four samples; the structural collapse deposit (Layers IIIa and
IIIb) yielded nine samples; and the basal diagenetic deposit yielded three
samples. Avian bones from the site were dated and boundaries between three
depositional units can be assigned absolute dates (Section 5.1).
Figure 7: Native, extinct land snail taxa at site 1710-1. Top, relative proportion with 1 errors; second from top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of G. servilis (note logarithmic scale). L. gracilis was not recovered from this site Analyse the data using XLisp-Stat
Collections from the basal diagenetic deposit are
characterised by an abundance of the aquatic snail, A.
nitida, and this accounts for the low relative proportion of native,
extinct taxa in these collections (Fig. 7, top). Davis believed that the sinkhole was at least
partially flooded throughout the period represented by this unit
(Davis 1990, 200), and basal diagenetic deposit collections strongly resemble
collections from low-lying, 'aquatic' sinkholes. The structural collapse
deposit marks the beginning of terrestrial land snail dominance in deposits,
but native, extinct taxa never dominate collections here as they do in older
collections of other sinkholes. Native, extinct taxa make up between 35±2% and 51±2% of the
structural collapse deposit collections, with a small, but significant dip in
the top two collections. The population of native, extinct taxa does undergo a
change at the boundary of transported sediments, where their relative
proportion drops to approximately 22% of the collection. It remains at this
level to the top of the stratigraphic column.
Diversity measures indicate that native, extinct taxa were
not under stress during the period represented by basal diagenetic and
structural collapse deposits. Both diversity measures rise slightly over this
time. The transition to transported sediments saw a marked decline in
diversity, with both measures showing a significant drop from previous levels.
The decline in diversity occurs in either the late
pre-Contact or post-Contact periods; 14C dating does not distinguish
between these possibilities (Fig. 3).
Distributions of introduced snails are also little help in this regard. The
Polynesian introduction L. gracilis is absent
from the site. The occurrence in small numbers of historically introduced
G. servilis in most structural collapse deposit
collections is almost certainly a result of mixing, and gives some indication
of the potential magnitude of this problem. The taxon is present throughout
the transported sediment deposit and this might be interpreted as an
indication that these deposits are post-Contact in age. Although absolute
numbers of this taxon rise a bit in the lower two collections from the
transported sediment deposit, their relative proportion does not increase
until the top two collections. Thus, an argument could also be made that G. servilis shells found below the top two collections
of the transported sediment deposit are intrusive.
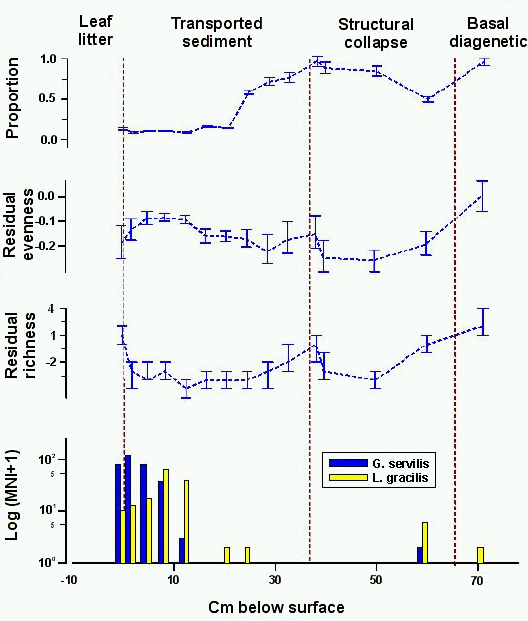
5.2.5 Site 9647-2
Excavations at site 9647-2 revealed 65cm of deposit in
four layers beneath a cobble fill (Davis 1995). The transported sediment
deposits (Layers II, III, IVa, and a portion of Layer IVb)
yielded nine
sediment samples for land snail identification; four samples were taken from
structural collapse deposits (Layer IVc and a portion of Layer IVb);
and a single
sample was collected from the basal diagenetic deposit (Layer V) (this sample was taken adjacent to the sample column (Christensen 1995, 243)). (The layer designations used here are those reported by Davis (1995) and by
Allen (1995). They designate rock fill at the site as Layer I; the first sediment deposit
is Layer II. Layer designations used by Christensen (1995) ignore rock fill and begin
with Layer I at the top of the sediment deposit. The Layer IVb sample was taken adjacent to the sample column
(Christensen 1995, 243).) A sample was
also taken from leaf litter at the surface of the deposit.
Collections from site 9647-2 reveal two periods of rapid
decline in relative proportion of native, extinct taxa. At the base of the
stratigraphic column, native, extinct taxa make up 96±4% of the collection from the basal diagenetic deposit
(Fig. 8). This is followed by a large drop to 50±3% at the base of the structural collapse deposit. The
structural collapse deposit is characterised by increases in relative
proportion of native, extinct taxa, which reach 97±6% at the top of the unit. Relative proportions
decline slowly through the bottom three collections in the transported
sediment deposits. The collection from 22-24cm below surface yielded 59±3% native, extinct taxa, a proportion slightly greater
than the collection from the base of the structural collapse deposit. The
second period of rapid decline brings the proportion to 14±1%, and it remains in the range 7-17% through to the
top of the stratigraphic column and into the leaf litter.
The first period of rapid decline in relative proportion of
native, extinct taxa is mirrored by a decline in diversity. The collection of
native, extinct taxa from the basal diagenetic deposit is the most diverse
from the site. Richness and evenness both decline significantly through the
transition from basal diagenetic to structural collapse deposits. The second
period of rapid decline in relative proportion of native, extinct taxa is
accompanied by a slight drop in richness, but this is barely detectable with
the materials and techniques at hand. Both diversity indices indicate little
change through the transported sediment deposit, which yields values of the
indices that are within ranges seen previously in structural collapse
deposits.
Figure 8: Native, extinct land snail taxa at sinkhole site 9647-2. Top, relative proportion with 1 errors; second from top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale). The sample from the basal diagenetic deposit was taken adjacent to the sampling column at a depth of 45-60cm below surface. This sample has been given an arbitrary depth of 70cm below surface to keep the deposits in their correct stratigraphic order
Analyse the data using XLisp-Stat
Under the assumption that depositional units are
penecontemporaneous among sinkholes, the first period of decline in the
population of native, extinct taxa at site 9647-2 took place in the early
centuries of the first millennium AD and the
second in the late pre-Contact or post-Contact period. The stratigraphic
distribution of the Polynesian introduction L. gracilis
appears to contradict the estimated age of the second period of
decline, however. Interpretation of the stratigraphic distribution of
introduced snails is not straightforward, and the collections yield evidence
for disturbance down to the base of the stratigraphic column. A single L. gracilis individual is present in the collection
from the basal diagenetic deposit (non-apical fragments of G. servilis were also found in this collection
(Christensen 1995, 240)), and it is found with G. servilis deep in structural collapse deposits. These
contexts are stratigraphically inferior to four collections in which both taxa
are absent. Single individuals of L. gracilis are
found midway through transported sediment deposits, but it is not until the
top of the stratigraphic column that either introduced taxon is found in
significant numbers. The mode of L. gracilis
distribution is earlier than that of G.
servilis, but both modes are later than the second period of decline in
relative proportion of native, extinct taxa at the site. If the assumption of
penecontemporaneity in depositional unit boundaries holds, then L. gracilis was not established at site 9647-2 until
very late in the pre-Contact period, or more likely the post-Contact period. (Allen suggests that the formation of the site 9647-2 sinkhole might have been
later than other sinkholes in the region, based on the low density of bird bones and
the apparent on-going structural collapse at the site (Allen 1995, 43). One
or two 14C dates from the base of the stratigraphic column would likely
provide the evidence needed to discriminate between this hypothesis and the
hypothesis that depositional regimes were penecontemporaneous throughout the
region.)
If the penecontemporaneity assumption is violated at site 9647-2, then all
that can be said is that the second period of decline pre-dated establishment
of an L. gracilis population at the site.
5.2.6: Site 9661-2
The sediment deposits of site 9661-2 were excavated in six
arbitrary 6cm and 10cm levels (Davis 1995, 331) (sediment deposits at site 9661-2 were capped by limestone rubble that was
excavated as Level 1 and assigned to Layer I). Six sediment levels were later
assigned to four natural layers and grouped into two depositional units. Basal
diagenetic and structural collapse deposits are present at the site, but
transported sediment deposits are absent. Seven samples for land snail
identification were recovered from the structural collapse deposit and a
single sample was taken from the basal diagenetic deposit. A sample was also
collected from leaf litter at the surface of the sediment deposit.
The relative proportion of native, extinct taxa declines
steadily from the base of the stratigraphic column, where they make up 76>±5% of the collection, to its top, where the proportion
is reduced to 9±1% (Fig. 9). The collection from leaf litter shows a slight,
though significant, increase in relative proportion to 20±4%.
Richness and evenness measures indicate a significant
decline in diversity of native, extinct land snail population near the top of
the structural collapse deposit. The population of native, extinct taxa never
regains its richness, but evenness recovers to previous levels by the top of
the deposit and maintains this level into the leaf litter. Fluctuations in
diversity in the early portion of the sequence are insignificant.
Figure 9: Native, extinct land snail taxa at site 9661-2. Top, relative proportion with 1 errors; second from
top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale). The site lacks a transported sediment deposit. Analyse the data using XLisp-Stat
Both introduced land snails are found throughout the
structural collapse deposit, and a few individuals of G.
servilis were recovered from the basal diagenetic deposit. If
depositional units at this site were correctly identified, then large numbers
of G. servilis in the structural collapse deposit
indicate substantial mixing of the stratigraphic column
(cf. Christensen 1995, 293). Otherwise, occurrence of G.
servilis through most of the structural collapse deposit indicates a
post-Contact age for the upper boundary of the depositional unit, and a very
recent age for decline in richness of native, extinct taxa at the site.
Although the timing of events at site 9661-2 is problematic,
the site shows a common pattern of change in native, extinct land snails.
Against a background of steady decline in their relative proportion beginning
in the basal diagenetic deposit, diversity indices remain stable until the
recent end of the sequence. Decline in diversity at site 9661-2 takes place at
a date that is likely to fall in the post-Contact period, if large numbers of
G. servilis are any indication of age, but the
decline cannot be dated with sufficient precision to rule out a date in the
pre-Contact era.
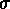 errors; second from top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale)
errors; second from top, difference of observed and expected evenness with 80% confidence intervals; third from top, difference of observed and expected richness with 80% confidence intervals; bottom, frequency of introduced taxa (note logarithmic scale)