Cite this as: Boismier, W.A., Allison, E., Ardis, C., Banerjea, R., Batchelor, C.R., Dark, P., Dudgeon, K., Green, C.P., Henderson, E., Ladocha, J., Weinstock, J., Young, D.S. and Schwenninger, J.-L. 2024 Investigation of Borrow Pit TEA28 BP3, Fenstanton, Cambridgeshire, UK, Internet Archaeology 67. https://doi.org/10.11141/ia.67.23
As part of the A14 Cambridge to Huntingdon road improvement scheme, Borrow Pit TEA28 BP3 was excavated in terrace deposits of the River Great Ouse to provide aggregate for construction works (NGR TL 3020 6779). These deposits were known to contain archaeological and environmental evidence of early human settlement, ancient landscapes and climate change spanning the Late Pleistocene period, 127-11.4ky (MIS 5e-MIS 2). Period resource assessments for the Palaeolithic set out in national and regional research frameworks have identified both the importance and rarity of this settlement evidence and its environmental context (Austin 1997, 2000; McNabb 2006; Pettitt et al. 2007; Medlycott 2011). Cambridgeshire County Council's Historic Environment Team (CHET), as a result of known Palaeolithic findspots, requested that an archaeological watching brief be undertaken during extraction operations as a mitigation strategy to identify and record Late Pleistocene deposits and to recover any archaeological and palaeoenvironmental materials associated with them. This report summarises the results of the work on the Late Pleistocene deposits at TEA28 BP3.
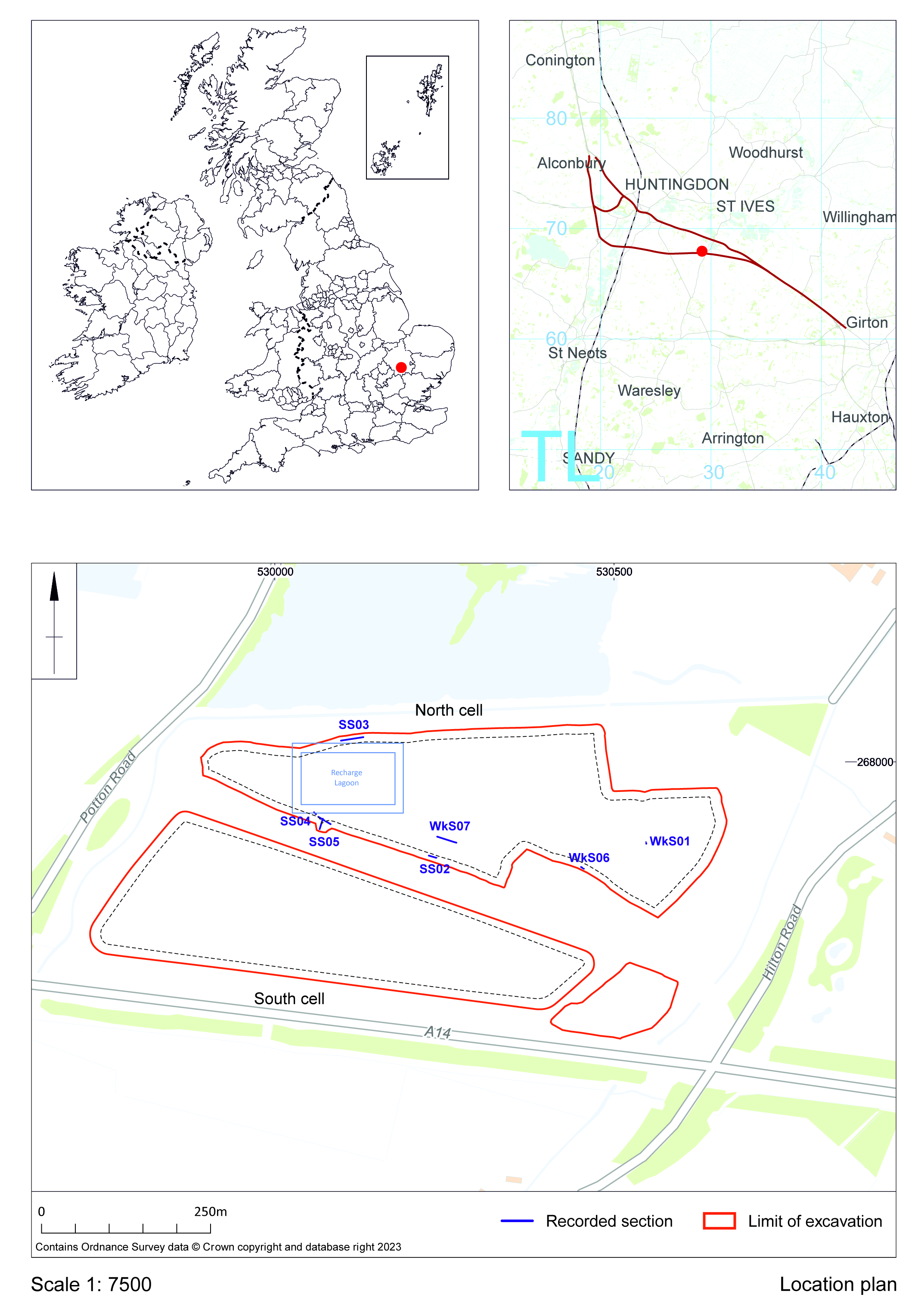
Borrow Pit TEA28 BP3 was located within an area of former gravel workings around Fenstanton and Hemingford Grey south-southwest of St Ives. In plan, the extraction area was roughly rectangular in shape and covered c.50.65 ha, and was situated to the south of the current A14 carriageway near Fenstanton, north-west of Cambridge (Figure 1; NGR TL 3020 6779). It was bounded to the north by flooded former quarry workings at West End Farm, to the east by Hilton Road, to the south by Hilton village, and along the western boundary by the B1040 St Ives road. The ground surface for the area has been mapped as lying between 7.1 and 8.3m above Ordnance Datum (aOD) (Gatfliff 1981). A brook and tree belt also ran along the north boundary, with additional trees present on the west and north-east sides. Former land-use was arable.
The borrow pit was situated on Quaternary river terrace deposits, mapped as the combined first and second terrace (T1-T2) of the River Great Ouse by the British Geological Survey (Edmonds and Dinham 1965; Gatliff 1981; Maddy 1999). These deposits comprise cold-stage river terrace sand and gravel of Middle-Late Devensian age (MIS 3-2; c. 60-15.4 ky), resting unconformably on Oxford Clay bedrock and covering an area of 11.1 km2 south of the River Great Ouse. Temperate deposits underlying these sands and gravels and dated to the Ipswichian Interglacial (MIS 5e; c. 127-115 ky) are recorded for nearby Woolpack Farm (Gao et al. 2000), Galley Hill (Cambridge 1964, 1965; Forbes and Cambridge 1966; Preece and Ventris 1983) and Somersham (West et al. 1999; Boismier et al. 2019). Similar organic channel-fill sediments with plant macrofossils and shell debris underlying third terrace (T3) deposits have also been found in two borehole logs from the Fenstanton-Fen Drayton area lying c. 1.4-2.3 km to the east-northeast of the borrow pit (Gatliff 1981, 67-68). Similar temperate deposits have not been recorded beneath river terrace sands and gravels within the area of TEA28 BP3.
The Palaeolithic archaeological record for the Fen Drayton-Hemingford Gray area (Figure 2) largely comprises isolated findspots and small collections of artefacts from quarries and other localities situated on river terrace deposits (Roe 1968; Wessex Archaeology 1996; Wymer 1999; Reynolds 1998, 2000; White et al. 2008; Cambridgeshire Historic Environment Record (CHER).
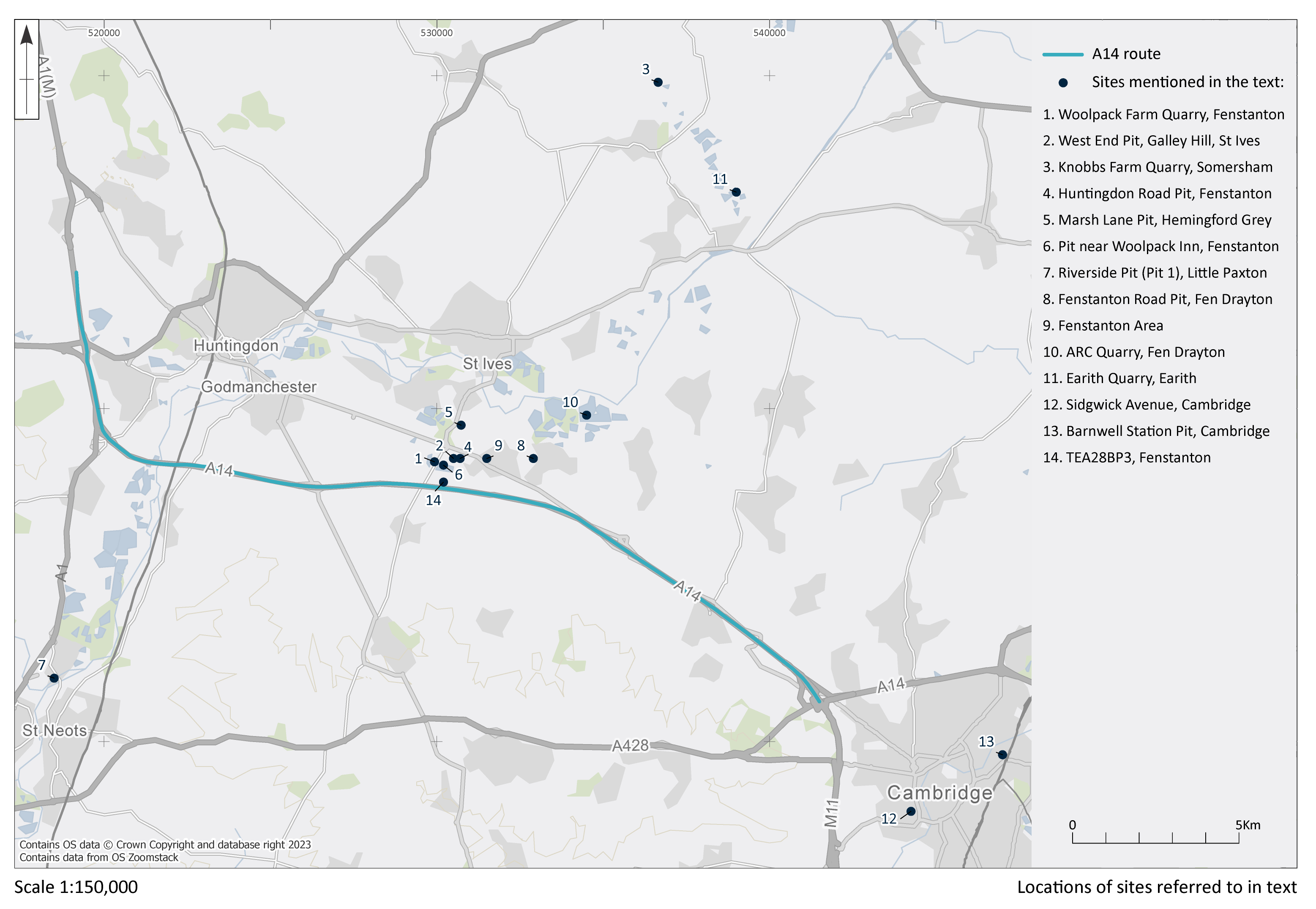
These include three handaxes, two of which are flat-butted bout coupé types of late Middle Palaeolithic date (MIS 3; c. 60-44 ky), and Levallois cores and flakes dating to the early Middle Palaeolithic (MIS 9-6; 300-186 ky), from former quarry areas situated on T1-T2 terrace deposits (CHER nos. 01692, 01693, TL302683). An important late Middle Palaeolithic artefact collection was recovered from T1-T2 terrace deposits at Little Paxton (CHER 0057) and includes 10-11 handaxes with at least one bout coupé type handaxe, handaxe manufacturing debitage and a number of flake tools. Former quarries on the T3 terrace in the Fen Drayton area have also produced collections of Lower Palaeolithic handaxes and early Middle Palaeolithic Levallois material (CHER nos. 01694, 03450, 10824). The recorded condition of this artefactual material ranges from severely rolled or abraded to unrolled or near 'mint' condition.
The monitoring of extraction works at TEA28 was undertaken to identify any in situ archaeological and/or palaeoenvironmental deposits exposed during these operations, and to record the succession of Quaternary deposits found at the site. This monitoring was carried out over 14 months between June 2018 and August 2019. Extraction excavations within this period proceeded from south to north, with the south cell of the quarry pit completed first and followed by the north cell and south-east corner of the site. On-site recording and collection activities ceased on 29 August 2019, when extraction excavation and backfilling for the north cell was completed and monitoring no longer required.
On-site work involved the recording and sampling of exposed quarry faces and the examination of surfaces and sections in working areas for in situ artefacts and vertebrate remains. Aggregate stockpiles and reject heaps were also examined for these materials throughout the period of site work. Detailed recording was carried out at seven selected locations in the north cell (Figure 1), on the north and south sides of the excavation area (SS02–SS05) and along working sections within the area (WkS01, WkS06–07). These locations were chosen on the basis of how representative they were in regard to depositional environments and lithological sequences across the extraction area. The recording for these sample sections included deposit and sediment descriptions, with representative sections and sedimentary features digitally photographed and located three-dimensionally with a global positioning system (GPS) (Figures 3–9). Samples were also taken from selected deposits in these sections, and included environmental (bulk, plant macrofossil, pollen), particle size, micromorphology and radiometric dating (OSL, AMS).
South cell deposits were less well recorded because of the rapid pace of excavation and backfilling operations. Records for this area largely comprise some deposit descriptions and digital section photographs for the north side of the extraction area, and a few working sections from within it and for the excavated south-east corner of the site. These are held in archive and available for further study.
The stratigraphic sequence for the north cell area was investigated utilising seven section samples with detailed recording. Section photographs, photo mosaics and on-site context sediment descriptions were employed to identify groups of environmentally related lithofacies that occurred together (facies associations) and their stratigraphic sequence across the extraction area (facies sequences). A sequence diagram (Harris matrix) was utilised in the analysis for facies grouping and stratigraphic ordering (Harris 1979; Goldberg and Macphail 2006, 40–41). OSL age estimates were also employed in the analysis to provide a dated time–stratigraphic framework for the sequence.
The analysis grouped the stratigraphic succession into four principal facies associations, which formed a sequence of fluvial, floodplain and periglacial colluvial deposits. These deposits were characterised by distinctive sets of lithofacies and stratigraphically separated from each other either by unconformities in the sedimentary record or by changes in depositional environments. This stratigraphic sequence is summarised in Table 1 and descriptions of the facies associations making up the succession are given below.
| Association | Facies Code | Facies | Sedimentary Structures | Interpretation |
|---|---|---|---|---|
| D | DMS | Matrix-supported massive sandy gravel; medium sand with fine-coarse flint clasts and rounded chalk fragments | Cryoturbation features (involutions) | Periglacial slope deposits |
| C | Fl | Silt, fine-medium sand | Horizontal bedded silt, sand and fine-small rounded gravel; cryoturbation features; convolute deformation bedding | Overbank floodplain deposits |
| B | Fm, Fl, P, Sr, Sp | Mud, organic, palaeosol, silt, fine-medium sand and gravel | Wavy and horizontal beds; fine lamination with interbedded sand, silt and organic sediments, rootlet horizon microfeatures (Fr); pedogenic (load structures) and cryoturbation features; convolute deformation bedding | Floodplain deposits: pond, marsh-fen, palaeosol sediments and stream or flood deposits |
| A | Gm, Sp | Massive or crudely bedded gravel, pebbly medium-coarse sand | Horizontal bedding, planar cross-beds | Longitudinal and transverse bars, lag deposits |
| - | - | Oxford Clay | - | Bedrock |
This association comprised sand and gravel lithofacies of fluvial origin and rested unconformably on fossilferous dark grey–blue Oxford Clay. Association A was dominated by sandy gravel lithofacies made up of massive and crudely bedded gravel, with some sand facies also present, and in places interbedded with and underlying gravel bedforms. Typically matrix supported, stony (very common–abundant 30–50%), with lithology dominated by fine–medium, rounded flint and lesser amounts of quartz, quartzite, sandstone, chalk and ironstone. The association was 0.30-2.5m thick and made up of short sequences of gravel and sand facies forming longitudinal bars and sheets similar to those recorded for gravel–sand-dominated braided rivers (Knighton 1998; Miall 2006). Silt and organic mud drapes also occurred, suggesting periods of low or slowly flowing water and low-stage subaerial exposure. The association formed a continuous basal deposit across the extraction area and was visible on vertical faces in excavation works and as patches of in situ sand and gravel covering clay bedrock.
Association B consisted of sand, silt and organic facies making up floodplain deposits and laid on the gravels and sands of Association A. It formed a laterally continuous deposit across the excavation area and was visible on vertical section faces and as excavated surfaces during extraction works. The association was c. 0.5-2.0m thick and characterised by the presence of local sequences on seven sections made up of organic sediments, including muds and palaeosols interbedded with fluvial sands and silts. These deposits are summarised below and as part of Table 1.
Facies for Association B on this section comprised deposits of planar and wavy–parallel medium–thick beds (0.11–0.39 m) of medium to coarse quartz sand–silty sand. These sands were interbedded with beds (0.04–0.17m) and laminae (3–10 mm) of sandy silt and organic sandy silt and clay. Sedimentary features included ripple and erosive ripple bedforms with inclined foreset laminae, and silt and organic mud drapes covering bedform crests and troughs along upper contacts (flaser-type bedding). Load cast deformation structures were also present in places along lower contacts. These beds suggested deposition from suspension and weak currents, with the succession of alternating sand and silt beds indicating repeated inundation from overbank flood events and likely periods of standing water.

Facies assigned to Association B for section SS02 comprised a thick planar bed (1.0 m) of bluish grey silt and organic sandy silt interbedded with fine–medium grained quartz sand–silty sand. The upper 0.67m of this deposit was extensively disturbed and displayed a series of irregular cryoturbation deformation structures and weathered (oxidised) sand. Lenticular platy ped microstructures within the thin-section for the deposit also provided evidence for cryoturbation and frozen ground. Sedimentary features in the lower 0.33m part of the deposit were better preserved and included interbedded parallel thin–medium beds (0.04–0.20 m) of sand, silt and organic sandy silt, ripple-like sandy bedforms and iron–manganese bands. These features and the fine-grained make-up of the sand, silt and organic sediments indicated that the bed represented a floodplain pond and/or marsh–fen deposit. Palaeobiological evidence (pollen, plant macros, insects) also suggested the presence of a shallow floodplain pond surrounded by sedge marsh–fens at the section locale.

Association B facies for section SS03 consisted of one thick planar bed ( 0.61 m) of medium to coarse quartz sand, with wavy–irregular medium beds (0.16–0.26 m) and laminae (3–7 mm) of organic sandy silt and mud. Cryoturbation features were present on part of this section and included ice-wedge cast, frost-shattered sediment blocks and weathered (oxidised) sand. Cryoturbation deformation structures were also suggested by irregular–contorted beds of organic sediment. Elsewhere, sedimentary features largely comprised silt and organic mud drapes covering sandy bedform crests and troughs (flaser-type bedding). These beds suggested alternating periods of flowing and standing water, possibly related to overbank flooding, with sand deposited during flow events and silt–organic muds by suspension during periods of standing water and weak currents.

No Association B sediments survived in situ for this section. Sediments were removed by machine during sump excavation prior to recording.

Association B facies comprised medium to thick beds (0.15–1.22 m) of organic and sandy sediments, and formed a succession of lower and upper organic beds that represented floodplain pond and/or marsh–fen deposits, stratigraphically separated by current-bedded sands representing stream or flood deposits. At the base of this succession an organic silty clay mud drape rested on the underlying gravels of Association A and was overlain by a deposit of gravelly sand with traces of horizontally bedded fine–small rounded gravel. The lower organic unit succeeded this deposit and made up a medium–thick bed (0.21–0.50 m) of silt, organic silty clay, and medium to coarse sand with fine–small gravel. The bed displayed a weathered appearance, with sedimentary features dominated by load cast deformation structures, and indicated a degree of sediment reworking by vertical displacement from sediment loading, and the forming of ball and pillow deformation structures with sand and gravel fills. Sedimentary structures were better preserved and more discernible in the upper bed and included wavy–irregular and parallel thin–medium beds (0.04–0.20 m) of sand, silt and organic sandy silt. Irregular–contorted and convolute sediment beds were also present, suggesting sediment deformation possibly related to cryoturbation. Current-bedded fluvial sand occurred stratigraphically between the two organic beds. It comprised a thick deposit (0.53–0.83 m) of horizontal and wavy bedded medium to coarse sand, gravelly sand and organic mud drapes. Sedimentary features included a succession of thin–medium beds (0.03–0.15 m) of sand and gravel with inclined foreset laminae, and fining-upward grading indicative of a gradual reduction in flow energy and an increase in suspension sedimentation. Palaeoenvironmental proxies (pollen, plant macros, insects) also suggested the presence of a shallow floodplain pond surrounded by sedge marsh–fens at the locale.

Facies for Association B comprised a palaeosol horizon and formed a vertical accretion deposit with a series of medium to thick beds (0.16–0.35 m) of organic and sandy sediments, suggesting several phases of soil development with periodic inundation and sediment deposition by overbank flooding. A thick bed of silty sand (1.11 m) lay at the base of this succession and was overlain by a sequence of medium–thick beds (0.16–0.35 m) of organic silty sand and medium sand–silty sand. Sedimentary structures for these facies included wavy–irregular thin–medium layers (0.05–0.26 m) and laminae (2–6 mm) of silt and organic silty sand. Traces of foreset laminae suggestive of low water velocity and weak currents were also present in sandy layers. Cryoturbation features on this section included ice-wedge casts, diapiric-type involutions and irregular–contorted beds of organic sediment, and weathered (oxidised) sand. Thin-section microfeatures for these sediments included traces of former soil and rootlet horizons, and microstructures indicative of cryoturbation. Palaeobiological evidence (pollen, plant macros, insects) also suggested the presence of a marginal wet–damp grassy meadow at the locale.

Palaeosol sediments also made-up Association B deposits in this section and were similar in make-up to those recorded in section WkS06. These sediments formed a deposit with medium–thick beds (0.17–0.38 m) of interbedded organic and sandy facies, again suggesting more than one phase of soil development. Sedimentary features for these deposits included wavy–irregular thin–medium beds (0.05–0.24 m) of organic silty sand, silt and sand–gravelly sand. Cryoturbation-like deformation structures were also present in places and comprised irregular–contorted sediment beds. Microfeatures on the deposit thin-section provided additional pedological evidence for soil freezing and cryoturbation.

This association comprised deposits that were laid down by overbank flooding on contact surfaces for Association B sediments. Association C was c. 1.50m thick, with a calcareous bluish grey to light brown silty clay matrix, and characterised by faint and finely laminated thin–medium horizontal beds (0.01–0.2m) of silt, sand, organic matter and fine–small rounded gravel. These beds suggested deposition from suspension and weak currents. Convolute bedding was also present along lower contacts, suggesting rapid increases in current velocity and flow turbulence possibly related to sheet flood events and/or the reactivation of an abandoned channel.
Sediments for this association were extensively weathered and disturbed by cryoturbation, with large parts of the deposit eroded and replaced by cryoturbation features and medium–coarse sand and gravel. These features included involutions, ice-wedge casts and frost cracks that extended from the overlying solifluction diamict into the deposit. Recorded redoximorphic features comprised weathered reddish yellow periglacial feature fills and light reddish brown silty clay mottling on a number of section faces across the extraction area (SS02, SS03, SS04, SS05).
The association survived as a series of isolated deposit fragments across the excavation area and was visible in places on vertical sections, quarry faces and working surfaces during extraction operations. These fragments suggested a fan-shaped deposit running in an irregular arc from north to east–southeast across the extraction area.
Association D comprised periglacial solifluction diamict (head) that rested on and partially eroded the silts of Association C. These deposits were typically c.0.59–1.0m thick and made up of calcareous pale–light brown and reddish yellow medium sand. They were stony (common–abundant 25–40%), matrix supported, and poorly sorted. Clasts were predominately fine–medium graded (3–16 mm), angular–subrounded flint and chalk gravel derived from the weathering of chalky till on nearby hills (Green 2000). Sedimentary structures were dominated by cryoturbation features filled with medium–coarse sand and gravel that extended into underlying deposits. Cryoturbation features included involutions and frost cracks. Ditches and other archaeological features such as pits were also cut into the upper c. 0.30–0.40m of the deposit. The association formed a laterally extensive deposit that extended across the entire extraction works area and was visible on vertical section faces and as chalky surfaces during excavation.
Soils for the extraction area comprised typical argillic brown earths of the Sutton 1 Association and lay on the chalky gravelly sand of Association D. The deposit was between 0.30 and 0.40m thick across the extraction area and made up of brown–dark greyish brown sandy silty clay. It was massive and stony (moderate 10–15%), with fine–medium and some coarse rounded flint, quartzite and occasional fine–small rounded chalk. The soils were plough and machine disturbed, with truncated profiles and made ground.
Samples taken from selected deposits in sections SS03, SS04 and SS05 were dated using OSL methods. Results for seven of these samples were judged to be unreliable because of probable age estimation errors caused by partial bleaching, mixed grain populations and chemical weathering. Age estimates for the remaining eight samples were considered to be reliable and in agreement with the stratigraphic succession of deposits. The results for these samples provide limiting dates for each of the facies associations and are summarised below and as part of Table 7.
Association A: Sands from this association in sections SS04 and SS05 yielded OSL age estimates of 43495±3635 (X7465), 41295±3155 (X7446) and 38810±2440 (X7447). These estimates appear internally consistent and form a sequence of dates in line with the succession of deposits, providing a basal age of c. 43 ky for the gravelly sands resting on clay bedrock and a date of c. 39 ky for upper contact sands.
Association B: Three OSL age estimates, of 40435±2845 (X7462), 40900±900 (X74630) and 41810±4400(X7449), were obtained for this association from fluvial sand interbedded with silt and organic facies in sections SS03 and SS05. All three of these estimates appear to be consistent with the stratigraphic record at around c. 41 ky and a Middle Devensian age.
Association C: Two age estimates were obtained from samples cut through redoximorphic features associated with cryoturbation structures in section SS05. Both were considered to be unreliable because of probable age overestimation errors from chemical weathering. OSL age estimates of 40435±2845 (X7462) and 22570±1570 (X7457) from the deposits sandwiching Association C constrain the age of this association and indicate that these deposits span the Middle–Late Devensian period and the transition to the last glacial maximum at around c. 27 ky (MIS 3–2; c. 60–15.4 ky).
Association D: An age estimate of 22570±1570 (X7457) for the chalky gravelly sand diamicton at the top of the succession dates it to within the last glacial maximum (MIS 2; c. 27–16 ky). A second date of 6895±540 (X7458) was also obtained for this deposit, from a sample taken immediately below the topsoil, and appears to be in good agreement with the stratigraphic record and associated post-glacial archaeological features cut into it.
Nine archaeological soil and sediment thin-sections were analysed to classify microstratigraphic units (MU) in order to understand and reconstruct this changing palaeochannel landscape (View report in digital archive [PDF]). MUs are classified into types of deposit and the origin of their components. To determine the deposit type classification, each deposit was grouped using the following diagnostic sedimentary attributes and inclusions, which provide information concerning the origin of inclusions, transportation mechanisms of particles and the deposition processes.
Soil micromorphology samples were prepared by Terrascope Thin Section Slides Laboratory, Troyes, France. To ascertain the origin of sediment components, descriptions were made of particle size and shape, and the composition of the coarse and fine fractions, particularly the frequency of rock, minerals and anthropogenic inclusions. The depositional events were characterised by the following sedimentary attributes: sorting, related distribution, orientation and distribution of the inclusions, and microstructure. Analysis of post-depositional features provides key information concerning the effects of weathering, preservation conditions (Stoops et al. 2018) and the stratigraphic integrity of the deposit (Kooistra and Pulleman 2018). Micromorphological investigation was carried out using a Leica DMLP polarising microscope at magnifications of ×40–×400 under plane polarised light (PPL), crossed polarised light (XPL) and, where appropriate, oblique incident light (OIL). Thin-section description was conducted using the identification and quantification criteria set out by Bullock et al. (1985) and Stoops (2003), with reference to Mackenzie and Adams (1994) and Mackenzie and Guilford (1980) for rock and mineral identification, and Stoops et al. (2018) for further identification of post-depositional alterations.
The micromorphological analysis produced several key findings.
Rootlet horizons are alluvial deposits that contain the abundant remains of roots, and probably reflect overbank deposition on a vertically accreting floodplain, such as discussed by Candy et al. (2006) in relation to rootlet beds. Eight MUs were identified as rootlet horizons, in all profiles analysed using soil micromorphology, except: MU2, sample 780007, context 782205; MU3 and MU5, sample 780046, context 782216; MU13 and MU18, sample 780059, context 782213; MU20, sample 780076, context 782212; and MU25, sample 780126, and MU28, sample 780128, both context 782403.
These MUs contained a dominant component of root and organic material (View report in digital archive, table 2 [PDF]), identified using Ismail-Meyer et al. (2018), comprising varying abundances of roots from mosses, the inner portions of roots, and amorphous organic material that was ferruginous or replaced with manganese. Fragments of leaves, seeds, vascular bundles and pseudomorphic plant voids, which are created where organic material has decayed in situ, also occurred in some of the rootlet horizons. The MUs, which varied in thickness from 5 to 42 mm, had a microlaminated bedding structure, whereby organic materials had rapidly accumulated and been compressed (Figure 10), unless, like MU2, they had been reworked by bioturbation, creating a massive bedding structure with the root material moderately oriented at a 45–90° angle to the basal boundary, rather than strongly oriented linear and parallel to the basal boundary (Figure 11).

MU5 has been classified as a stabilisation horizon and may also represent a rootlet horizon (Figure 11). It had a microlaminated bedding structure formed by gentle inundations of sediment interspersed with abundant root material (View report in digital archive, table 2 [PDF]) that was decomposing into the soil matrix.

Palaeosols, or buried land surfaces, were identified in samples from two sections: section WkS06 (context 782403, samples 780126, 780127, 780128) and section WkS07 (context 782505, sample 780138). Sample 780128 was collected from below sample 780127. It showed an earlier palaeosol/former B horizon, MU30, which was covered by a sequence of slow-flowing waterlaid inundations of sediment (MUs 27 and 29) and a rootlet horizon (MU28). Vesicles in MU29 could be indicative of standing water. Unlike MU24, the palaeosol/former B horizon MU30 was not covered by an overlaying A horizon, which may have been eroded away.
Sample 780127 (Figure 12) revealed a sequence of a soil profile, MUs 23 and 24, that was covered by a waterlaid inundation of sand, MU22, similar to MUs 4, 6 ,8, 16 and 21, which are reported above.

MU24 was substantially reworked and revealed the features of a relict soil with lenticular platey peds indicative of cryoturbation (Retallack 2001; Cremaschi et al. 2018). It is not uncommon for palaeosols to have been continuously reworked and to have several phases of bioturbation (French 2003). MU24 had two microfabrics (View report in digital archive, table 1 [PDF]): a and b. Decayed root inclusions (ibid. table 2) were associated with MFb. There were abundant infillings, 35% (ibid. table 3), of root channels infilled with silty clay, MFb, which was probably translocated from MU23 above. MU24 could be the former B horizon and MU23 the former A horizon. A large, c. 1cm, angular flint, 5%, was located at the boundary between MU23 and MU24 (View report in digital archive, table 2 [PDF]). This could be a stone tool, but further confirmation from the artefact assemblage is required. MU23 had a slightly different composition of organic materials to the other MUs, which were classified as rootlet horizons (View report in digital archive, table 2 [PDF]); conjoined elongated phytoliths, 2%, which form in the leaf and stem of grasses, were identified, and a fragment of a leaf, 3%. The bulk of the organic component, 40%, was amorphous.
Sample 780126 was collected from above sample 780127, and showed a continuation of the soil profile that was identified in sample 780127, with the caveat that this slide was very poorly prepared. MU25 was a rootlet horizon, and MU26, below, a former A horizon that shared similar sediment properties with MUs 23 and 32 and, again, the bulk of the organic component was amorphous, differing from the rootlet horizon, MU32, which contained fragments of the inner portions of roots, 15%.
Like MU24, MU30 was substantially reworked and showed the features of a relict soil or palaeosol (Retallack 2001). Also like MU24, MU30 had two microfabrics (View report in digital archive, table 1 [PDF]), a and b, as well as very abundant infillings, 30% (View report in digital archive, table 3 [PDF]). Framboidal pyrite had formed in MU30, <5%.
Sample 780138, field context 782506, comprised three MUs: waterlaid sediment–medium flow, MU31; a former A horizon, MU32; and a palaeosol/former B horizon, MU33. MU31 shared similar sediment. Only the upper 0.8cm of MU33 was included in the thin-section, and it was identified as a palaeosol/former B horizon by its oxidised orange/1st order red colour, PPL, infillings, 20%, which showed reworking, and an overlying A horizon, MU32. The A horizon contained fragments of a pellet, 15%, belonging to a small herbivore. It resembled the morphology and internal structure of a sheep or goat coprolite (Brönnimann et al. 2017) but, in this context, it is more likely to be from a small deer, the coprolites of which share morphological characteristics with those of sheep and goat (Bang and Dahlstrom 1974, 183). Flint inclusions in MU32 and MU33 were <1cm in size and subangular in shape. These inclusions may represent microdebitage (Fladmark 1982) that had been transported in flowing water, causing the rounding of the edges during transport.
Rootlet horizons and soil profiles had formed within a system of overbank inundations of alluvial sediment, which had been transported and deposited by varying speeds of flowing water, ranging from sand or loamy sand deposits that were moderately sorted, indicating a higher energy environment, compared with those with a greater silt or clay composition that had a well-sorted silt fraction, indicating slow-flowing or standing water, particularly where vesicles and vughs (collapsed vesicles) occurred in the microstructure (View report in digital archive, table 1 [PDF]). Vesicles can form where air or water droplets become trapped in the sediment. Notably, MUs 19 and 21 contained large, angular-shaped flints, which could be stone tools (Angelucci 2017). The flint inclusions within MU21 were probably redeposited by fast-flowing water, but those in MU19 may have been dropped into standing water.
Changes in the hydrological system could be seen in the range of post-depositional alterations that were identified. Both impregnative and depletion redoximorphic pedofeatures were present, evident by the dotted b-fabric (View report in digital archive, table 1 [PDF]) arising from the formation of iron and manganese nodules and iron coatings ((View report in digital archive, table 3 [PDF])), which can result from long periods of saturation and fluctuations in the water table (Vespraskas et al. 2018). Most MUs showed manganese replacement of organic materials, which occurs at the capillary fringe (Bartlett 1988), but it occurred most abundantly in rootlet horizons. Calcite-depletion coatings had formed in MU1 and MU20, whereby calcite from the sediment matrix had dissolved and reprecipitated around voids, which is probably again the result of a fluctuating water table or root activity (Candy et al. 2006; Durand et al. 2018). Framboidal pyrite had formed in several MUs, particularly rootlet horizons, but also in palaeosols. These framboids commonly occur in marine and perimarine environments and form at the oxic–anoxic interface (Kattenberg and Aalbersberg 2004; Mees and Stoops 2018), but also in freshwater environments as a result of the bacterial degradation of organic matter (Ismail Meyer et al. 2013). Gypsum dissolution features, 2–5%, were observed in voids within MUs 9, 10, 12, a3 and 16, which can be indicative of surface horizons, or seen at the capillary fringe in saline environments, or in calcareous sediments where there have been large volumes of water (such as in irrigation) in a Mediterranean climate (Poch et al. 2018). Several MUs had lenticular platey peds within the microstructure, which can form as a result of cryoturbation (Van Vliet-Lanoë and Fox 2018).
One serial column sample, 78003, was used to investigate the depositional environment for the succession of fluvial sands sandwiched between organic floodplain and overbank deposits on the east face of section SS05 (contexts 782212, 782211, 782213, 782206). Nine samples were taken through the succession of sediments in this section, and subsampled (c. 10 g) for particle size distribution analysis. Subsamples were prepared utilising established procedures for laser granulometry, with particles falling within the size range 0.01–2000 μm measured by laser granulometry using a Malven Mastersizer 3000. Clasts greater than 2000 μm (2 mm) in size were removed prior to the analysis (Young et al. 2020).
All subsamples formed a stratigraphic sequence and showed a 'fining upward' grading (Figure 13), with the proportion of sand decreasing from c. 85–95% in contexts 782212 and 782211, falling to c. 60–70% in context 782213 and then sharply to between 20 and 10% at the top of context 782213 and into 782206.
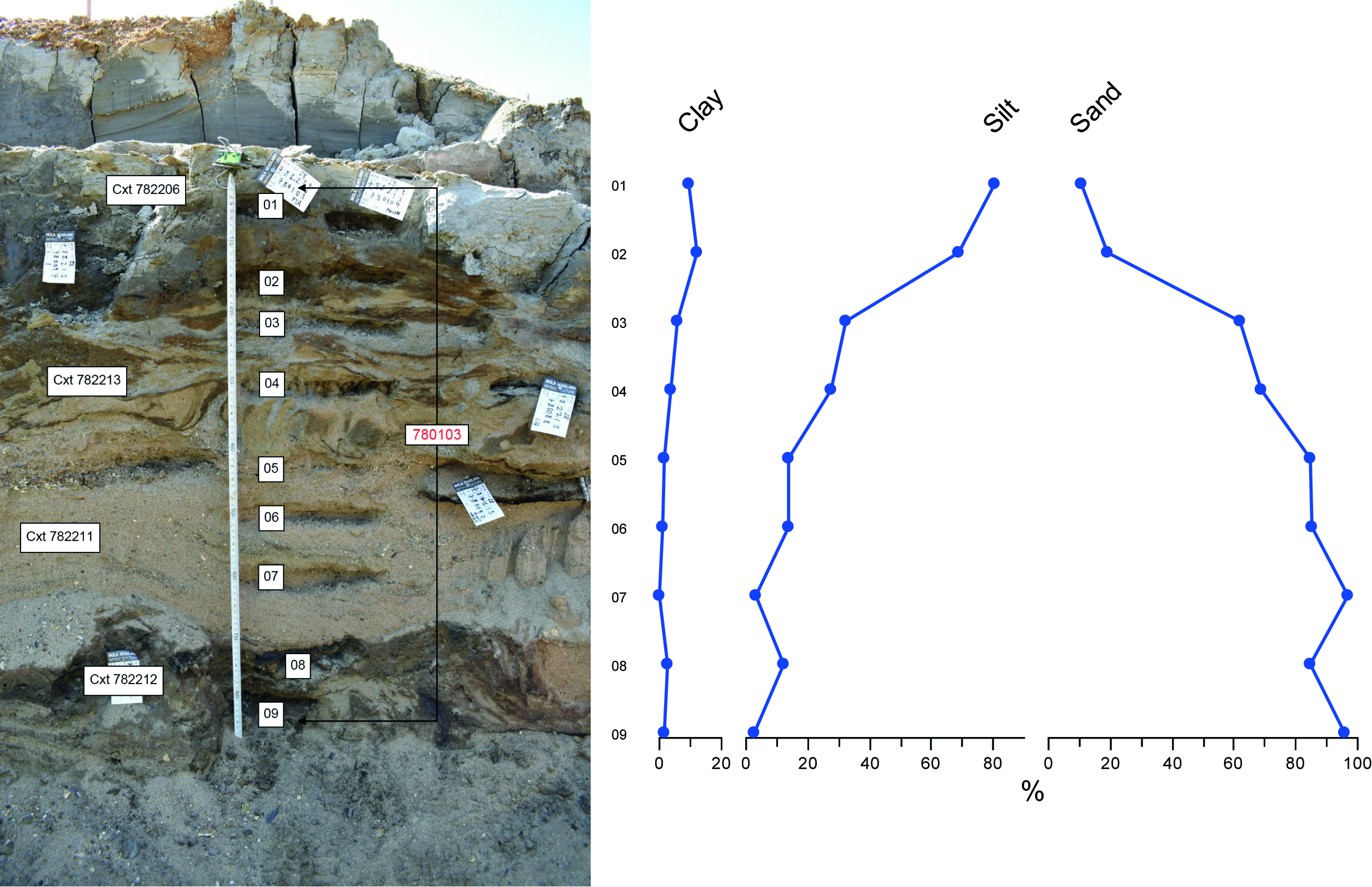
The proportion of silt rose, reflecting the reduction in sand content, with silt forming between c. 2 and 15% in contexts 782212 and 782211, rising to c. 70% in context 782213 and then 80% in context 782206. The proportion of clay remained low throughout the sequence (0–6% in contexts 782212, 782211 and 782213, but rising to c. 9–12% in context 782206. There was no indication in this sequence for sediments with an aeolian origin (loess), with no samples with modes in the coarse silt fraction (Figure 14).
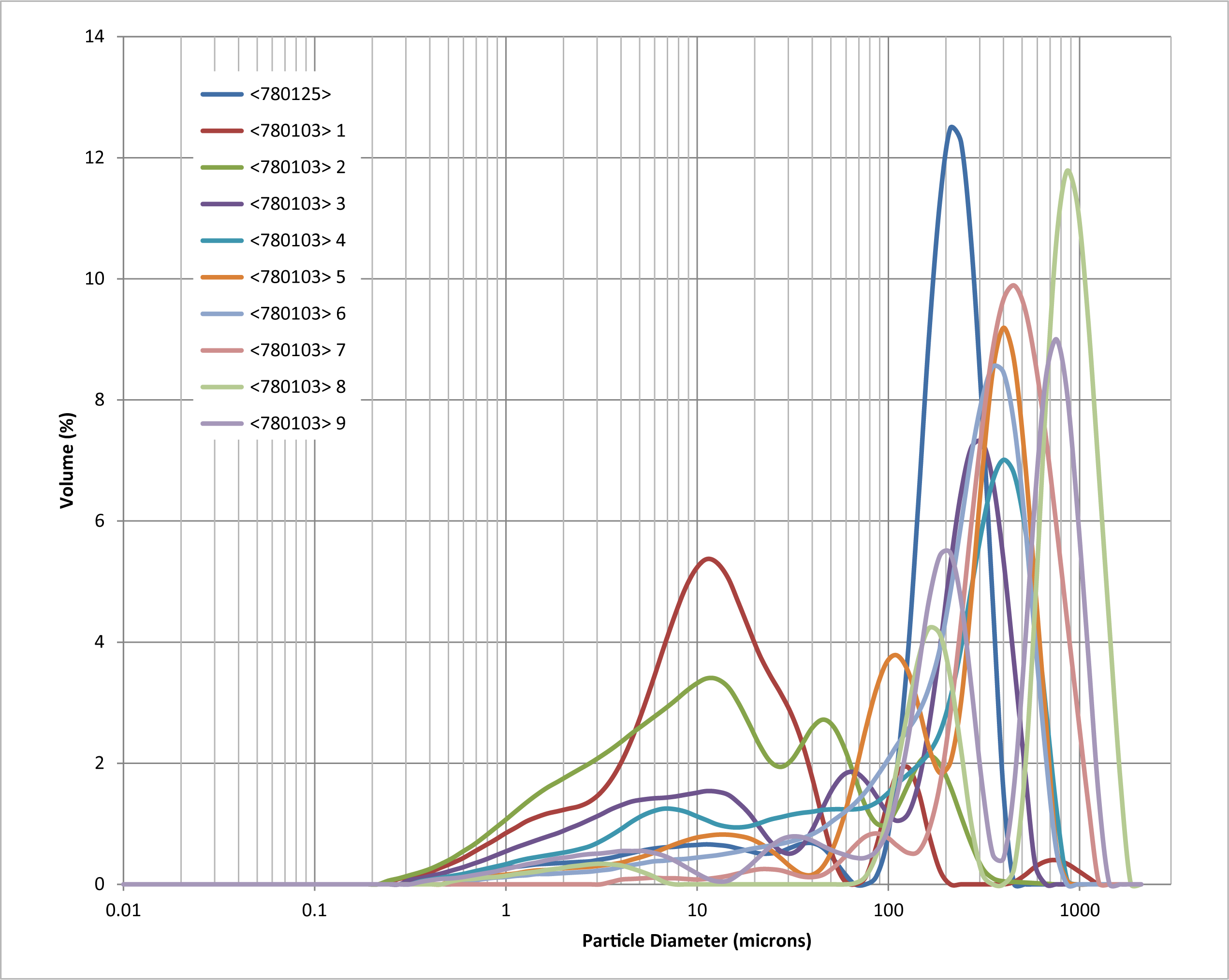
The PSD was consistent with fluvial deposition of the sediments, and indicative of a gradual reduction in flow energy through contexts 782212, 782211, 782213 and 782206. There was possible evidence for deposition of clay in suspension (indicative of standing water) in context 782206, although this context was at least partly periglacially disturbed, and the clay may be derived from post-depositional mixing of the sediments.
Analyses of pollen, micro-charcoal and parasite eggs were undertaken on two monoliths and a bulk sample from Late Pleistocene terrace deposits of the River Great Ouse revealed in Borrow Pit 3. OSL dates suggested that these deposits were of mid-Devensian age (MIS 3; c. 40 ky) (Boismier et al. 2021). An initial assessment of multiple samples (Young et al. 2020) identified five meriting further analysis, and these form the subject of this report (View report in digital archive [PDF]).
The monoliths 780059 and 780063 and bulk sample 780084 were from the south side of the excavated area, and all subsamples analysed here were from Association B. This comprised interbedded sand, silt and organic sediments, having formed in a floodplain environment with subsequent disturbance by cryoturbation. The monoliths were from contexts identified by micromorphological analysis (Young et al. 2020) as representing periods of relatively stable deposition, while the bulk sample was from an organic mud layer.
The samples were prepared using the standard technique of Quaternary Scientific, University of Reading (Branch et al. 2005), and a minimum of 300 identifiable pollen grains and Pteridophyte (fern) spores counted in each. The results were tabulated (Table 2 PDF) and taxa forming >1% of the sum displayed as percentage pollen diagrams (Figures 15, 18, 19).
Micro-charcoal particles and parasite eggs were counted at the same time as the pollen grains, and Lycopodium markers used to calculate concentrations for the former (Table 2 (PDF), Figures 15, 18,19).
It is important to consider the sources of pollen in deposits, key factors being the effects of differential preservation on the representation of some taxa, and reworking of earlier deposits (e.g. Lowe 1984).
Pollen preservation was highly variable, especially in the basal sample from monolith 780059. Many of the pollen grains in this sample were highly fragmented or crumpled, while others appeared as 'ghost grains', where details of the grain structure were no longer visible. In such cases identification was impossible, or possible only for the most distinctive pollen types, such as those of the Lactuceae, which remain recognisable even in a fragmentary state. The assemblages had thus been affected by both differential preservation, favouring taxa that are relatively resistant to deterioration, such as conifers, and differential identification, favouring those identifiable in an advanced state of deterioration.
Floodplain deposits frequently contain a high proportion of reworked and redeposited pollen, and reworking probably accounted at least in part for the poor preservation of much of the assemblages. The relatively abundant pollen of conifers in some samples, much of it too fragmentary to identify further, but including a significant proportion of Picea (spruce) but little Pinus (pine), suggests reworking of earlier Pleistocene sediments. Picea is generally recorded only sparsely from cold-stage assemblages, but was probably a significant component of Early Devensian interstadial forests (West 2000, 82).
Reworking was also suggested by the relative abundance of pre-Pleistocene palynomorphs, especially in the basal sample from monolith 780059. Much of the micro-charcoal in the samples may also represent reworking rather than contemporary fires, the highest charcoal frequencies occurring in this same sample.
Description
Pollen was abundant in all three samples, but preservation was variable (Table 2 (PDF), Figure 15). All samples contained some well-preserved pollen, such as Cyperaceae (sedges), presumably broadly contemporary with the period of deposition, in contrast with that likely to have been reworked (conifers especially). Pre-Pleistocene palynomorphs were abundant in the lowermost sample (c. 10%), declining in the two successive samples.
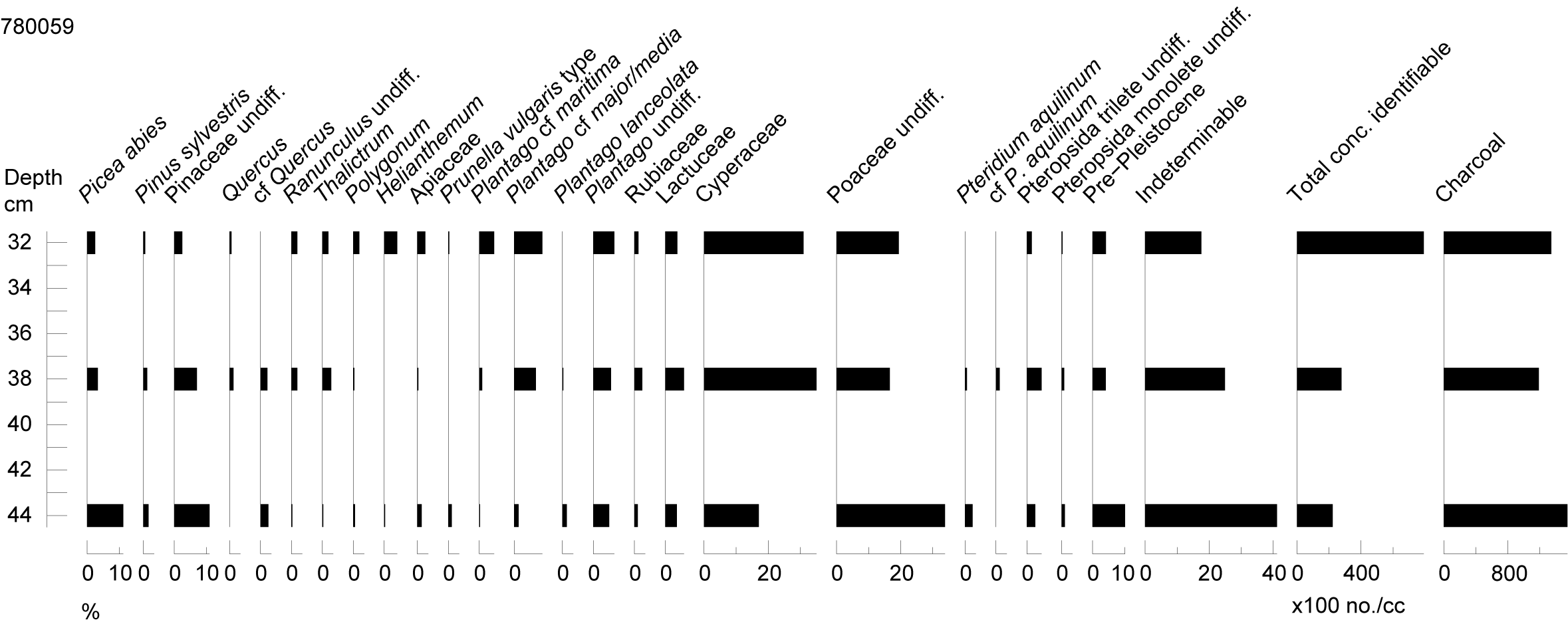
The pollen assemblages were dominated by herbaceous taxa, particularly Cyperaceae and Poaceae (grasses), with other herbs present, including Thalictrum (meadow rues) and Plantago (plantains). Poor preservation precluded specific identification of some of the Plantago grains, but P. maritima (sea plantain) and P. media (hoary plantain) and/or major (greater plantain) appeared to predominate. Pollen of Helianthemum (rockrose) comprised 4% of the uppermost sample, while rarer taxa of note included Limonium (sea lavender), Linum undiff. (flax) and Armeria maritima (thrift). Arboreal pollen consisted almost entirely of conifers, likely to represent a combination of pollen, Picea (spruce) especially, reworked from earlier Pleistocene deposits, and perhaps long-distance transport.
Some changes were apparent moving up the sequence, especially in terms of the relative abundance of grasses and sedges, but these probably reflected improved preservation (sedge pollen being relatively delicate, and difficult to identify when deteriorated) and a declining proportion of reworked pollen in the two upper samples.
Micro-charcoal was abundant in all samples, probably largely reworked from earlier deposits, as the highest values were associated with the greatest proportion of Picea, pre-Pleistocene and indeterminable pollen.
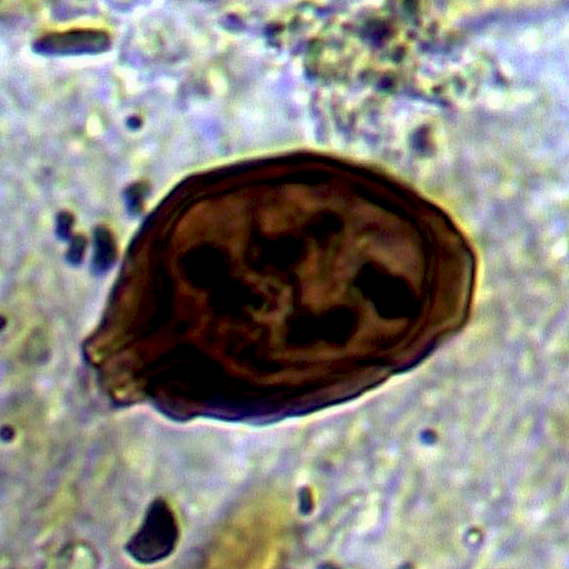
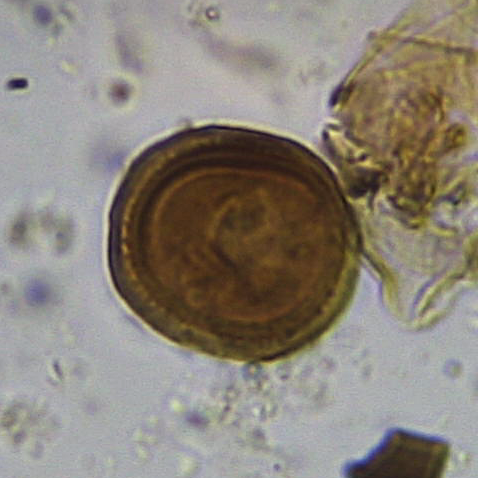
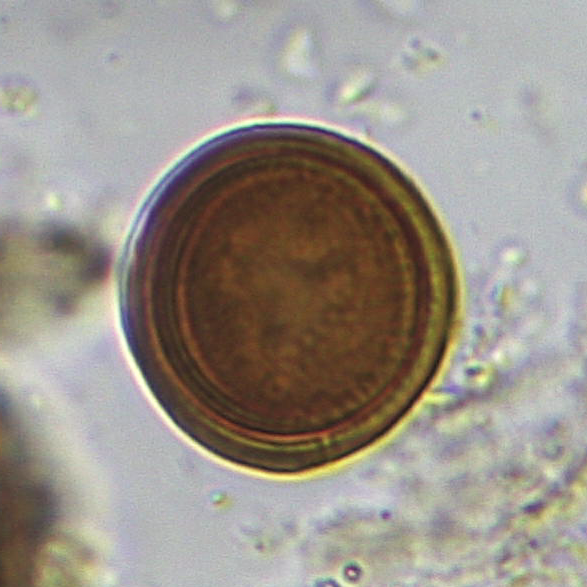
The sample from 44cm contained what appeared to be an egg of the intestinal helminth parasite Trichuris (whipworm) (Figure 16), as well as three eggs resembling those of a cestode (tapeworm) such as Taenia (Figure 17a, b). Four further cf. cestode eggs occurred in the sample from 38cm.
Interpretation
Overall, the assemblages suggest a very open environment on a floodplain, dominated by grasses and sedges, with a range of other herbs typical of cold-stage flora, such as Linum (flax) and Armeria maritima (thrift) (cf. West 2000). Areas of damp (including sedges) and drier [(with Helianthemum (rockrose) and Artemisia (mugwort)] grassland are indicated, while the presence of halophytic plants typically associated today with saline conditions in coastal settings, such as Limonium (sea lavender), Armeria maritima and Plantago maritima (sea plantain), may suggest locally saline conditions created by high evaporation rates in a strongly continental climate (West 2000, 168). This grassland environment would have been attractive to grazing herbivores, their dung presumably being the source of the intestinal parasite eggs present in the sediments. It is possible that grassland fires occurred periodically, although much of the micro-charcoal in the deposits was probably reworked.
Description
The single sample from this monolith contained a high concentration (c. 54,000 grains cm3 /cc) of pollen with a variable state of preservation, 16% being indeterminable (Table 2 (PDF), Figure 18).

The assemblage was dominated by pollen of Poaceae (grasses), accompanied by Cyperaceae (sedges) and Plantago spp. (plantains), with lower percentages of other herbs, including Thalictrum (meadow rues), Helianthemum (rockrose) and Artemisia type (mugworts). Interestingly, two large Poaceae grains, with maximum diameters of 47 and 58 μm, and annulus diameters of 12 and 9 μm, respectively, were recorded. Had they occurred in mid–late Holocene deposits they would have been thought likely to represent cereals, but in this context they must represent wild grasses with large pollen grains, such as Leymus arenarius (Lyme grass) (cf. Andersen 1979).
Arboreal pollen was confined to a low percentage of conifers (3% in total) and sparse records of deciduous taxa: Quercus (oak), cf. Populus (aspen) and Salix (willow).
Micro-charcoal was abundant (c. 126,000 particles/cc cm3 ), and a single egg resembling that of a cestode (tapeworm, cf. Taenia) parasite was present.
Interpretation
This sample resembles those from monolith 780059 in general terms, although with a lower representation of (presumably reworked) conifer pollen, coinciding with a smaller proportion of indeterminable and pre-Pleistocene pollen. An open grassland environment is indicated, with at least some areas probably grazed by large herbivores.
Description
This sample was notably different from the others examined. Preservation was better, with only c. 5% of the pollen unidentifiable as a result of deterioration, and pollen concentrations were relatively high, at c. 71,000 grains/cc (Table 2 (PDF), Figure 19). No pre-Pleistocene palynomorphs were encountered.
The most remarkable feature of this sample was the abundance of Centaurea scabiosa (greater knapweed) pollen, comprising 26% of the sum. Poaceae (grasses) and Cyperaceae (sedges) occurred, representing 9% and 13%, respectively, while species of Plantago (plantains) together reached 23%. Other herbaceous taxa included Thalictrum (meadow rues), Helianthemum (rockrose), Linum catharticum (purging flax) and Artemisia type (mugwort). Large Poaceae, all with maximum diameters of 55 μm or more, comprised 1% of the sum.
Arboreal pollen was barely present, while micro-charcoal was much less abundant than in any other sample, at c. 12,000 particles/cm3.
Interpretation
This sample represents a very open cold-stage environment of species-rich grassland, in which it seems that plantains and greater knapweed were significant components. It is possible, however, that knapweed is overrepresented because of the inclusion of part, or all, of a flower in the sample, although none of the pollen was in clumps, which may be the case in such situations.
The quantity of micro-charcoal in this sample was approximately an order of magnitude less than in any of the others, probably providing a more accurate representation of contemporary charcoal inputs than the other assemblages, which were clearly affected by reworking. It may be that the continental climate, inferred from the plant communities, encouraged burning of the floodplain vegetation in the summer months.
The pollen and spore assemblages from the Borrow Pit samples suggest cold-stage stadial conditions of open species-rich grassland with a variety of plant communities reflecting variable hydrological conditions on the floodplain, including areas of relatively high salinity supporting plants that would today be characteristic of coastal areas. Arboreal pollen consisted largely of Picea (spruce) and other conifers, almost certainly representing reworking due to their generally poor state of preservation and correlation with relatively high percentages of deteriorated pollen, pre-Pleistocene palynomorphs and micro-charcoal.
There are few comparable Middle Devensian sequences with adequate preservation of plant material, although macroscopic remains apparently of a similar date have been recovered from Sidgwick Avenue, Cambridge (Lambert et al. 1963), and Earith, Cambridgeshire (Bell 1970). At the latter site the plant remains were interpreted as representing glacial conditions, but coleopteran analyses (Coope 2000) suggested that there were two different climatic regimes, one with a cold continental climate, and a second, probably younger, with warmer summers, with mean July temperatures of around 16℃. Despite this relatively temperate climate, the contemporary landscape appears to have remained treeless. This demonstrates the extent to which plant communities may display a lagged response to climate change, and the necessity of using coleopteran evidence in preference to plant remains to infer past climate regimes.
An aspect of the Middle Devensian environment that has received little attention is the possible role of fire in influencing plant communities, probably due to the scarcity of combined pollen and micro-charcoal analyses. Although much of the micro-charcoal present in the borrow pit samples appears to have been reworked, there are indications that some burning was contemporary with deposition of the sediments, raising the possibility that fire may have been a significant factor in maintaining the diversity of habitats on and near the floodplain. Wetland and grassy vegetation may burn readily in the right conditions, as at Early Mesolithic Star Carr, Yorkshire, where most of the micro- and macroscopic charcoal was derived from reeds (Phragmites australis) (Dark 1998; Hather 1998). At Star Carr, a human source of ignition was likely because of the close association between the charcoal horizons and artefacts. Generally it is more difficult to distinguish between natural and anthropogenic fires. Archaeological evidence for human activity contemporary with deposition of the floodplain deposits at the borrow pit is slight, a single handaxe fragment that may be reworked (Boismier et al. 2021, 27), but it is nevertheless possible that human activity provided the source of ignition for at least some of the burning.
The occurrence of an egg of Trichuris, and possible cestode eggs, is of interest as records of intestinal parasites of this date are very rare. Most likely these derive from parasitic infections of large herbivores grazing the rich floodplain vegetation, although a human source should perhaps not be completely excluded. The OSL age estimates of 51285±3760 (X7451) and 45475±5130 (X7452) for the Association B deposits providing this evidence (contexts 782213, 782216) were considered unreliable because of estimation errors related to partial bleaching and the fluvial reworking of older sediments into younger deposits. An AMS radiocarbon date of >36300 (OxA-43556) for one deposit (context 783316) also lay beyond the reliable limits of the radiocarbon method, and indicated that the true age was probably in excess of 36 ky.
Possible cestode eggs of a similar date to those from the borrow pit have been recorded from mammoth remains from Russia (cited in Slepchenko and Reinhart 2018), but previous records of Trichuris appear to be confined to the Holocene (Gonçalves et al. 2003), the oldest examples in north-west Europe coming from Late Mesolithic intertidal peat deposits at Goldcliff, south Wales, directly associated with archaeological evidence for human activity (Dark 2004). Borrow Pit 3 deposits thus appear to provide the earliest evidence for whipworm infection in Europe, and possibly worldwide.
A total of 10 bulk samples was extracted and processed for the analysis of plant macrofossil remains, taken from facies Association B, c. 0.2–0.5m thick and comprising sequences of interbedded sand, silt and organic sediments indicative of alluvial deposition from overbank flooding and in situ soil formation (Boismier et al. 2021). A total of 1.5 litres of sample was processed for all but samples 780084 and 780042 (0.6 and 1.0 litres, respectively). The extraction process followed standard procedures for extracted waterlogged plant macrofossils, including wet sieving using 125 μm, 300 μm and 1 mm mesh sizes. Each sample was scanned under a stereozoom microscope at ×7–×45 magnification. Identifications were made using modern comparative material in the University of Reading reference collection, and reference atlases including Martin and Barkley (2000), NIAB (2004) and Cappers et al. (2006). Nomenclature follows Stace (2010). The taxa were arranged into broad habitat classifications to aid the interpretation of the assemblage and reconstruction of past vegetation and palaeoenvironment. The 'unclassified' group contained taxa that were at too high a taxonomic level to allow classification into a habitat group, or taxa that can be found in a number of habitat types.
The results of the plant macrofossil analysis, with the taxa arranged into broad habitat classifications, are shown in Table 3 (PDF), and Figure 20. The results of the analysis are summarised below by section sample (View report in digital archive [PDF]).

A single sample 780001 was analysed from section WkS01, context 782040, a coarse sand, silt and mud/carbonaceous mud. Plant macrofossil remains were very poorly preserved in this sample, with only a single fruitlet of Potentilla sp. (cinquefoil) and a seed of Linum cf. perenne (perennial flax) identified. This assemblage was too small to attempt an environmental reconstruction, but both species are typical of grassland habitats or grassy heaths.
One sample 780014 was analysed from section SS02, context 782205, an organic medium sand, silt and clay interpreted as a root bed. Plant macrofossils were well preserved in this sample; the most dominant components of the assemblage were aquatic, making up nearly half of the assemblage (n = 121), with others typical of waterside environments or damp ground or open grassland habitats. The presence of perennial flax, a coloniser of open habitats that requires high light intensity, is a typical component of Devensian palaeoenvironmental records (Bell 1970). Combined, the assemblage is indicative of deposits forming within a shallow water habitat such as a stream or pond, probably including sediments derived from fluvial input from the wider floodplain incorporating species from waterside and adjacent grassland environments.
A total of seven samples was analysed from section SS05, including contexts interpreted as root beds or organic muds, comprising organic fine to medium sands, silts and clays. Plant macrofossils were generally well-preserved in the samples from contexts 782212, 782213 and 782216, although preservation was comparatively poor in the samples from contexts 782228, 782231 and 782238. The plant macrofossil assemblages were generally dominated by aquatic taxa and those typical of waterside environments or damp ground, with some found within open grassland environments. Seeds of Cornus cf. sanguinea (common dogwood) were recorded in the samples from contexts 782213 and 782231, representing the only recorded shrub species with tree taxa entirely absent. This species requires light and alkaline soils and is often found in riverside environments.
Similar to the contexts from section SS02, the assemblage in these samples was indicative of deposits forming within a shallow water habitat such as a stream or pond, likely incorporating sediments derived from fluvial input from the wider floodplain and including species from waterside and adjacent grassland environments.
A single sample 780132 was analysed from section WkS06, context 782803, described as sand, fine gravel and mud and interpreted as a palaeosol. The assemblage was dominated by sedges, with some aquatic taxa. The assemblage was relatively small in this sample, but was indicative of waterside environments and damp ground likely adjacent to a pond or stream.
As a whole, the plant macrofossil assemblages from the samples are indicative of sediments forming in fresh, shallow, still or slowly flowing water, likely incorporating material from a wider floodplain, including taxa growing in waterside environments, damp ground and adjacent grassland. Although brackish or saline influences have been recorded in parts of the sequence elsewhere in deposits from the River Great Ouse, evidence for this is limited here to horned pondweed, which grows in a range of shallow-water habitats, including eutrophic and brackish environments. The macrofossil assemblage indicates that the sediments are likely to have incorporated material derived from fluvial redistribution of sediments from the wider area of the floodplain, including marginal environments close to the channels or ponds in which the sediments were deposited. The absence of finer plant macrofossils, for example leaves of mosses and grass caryopses, may be a result of hydrodynamic sorting in flowing water. However, on the basis of the preservation of the macrofossil remains, there is little evidence to suggest that there had been substantial reworking of the sediments, and the seeds are unlikely to have been transported far from their source area.
In general, the assemblage lacks any southern, thermophilous elements. In the absence of such taxa there is no conclusive evidence for warmer conditions associated with an interstadial climate. The data from palaeoenvironmental reconstructions for this period in Britain are inconclusive (see Buckland et al. 2019), and potentially complicated by poor chronological control and the reincorporation of material from older sediments in fluvial contexts. However, a highly variable environment likely incorporating climates typical of both stadial and interstadials has been suggested by Buckland et al. (2019). Plant macrofossil deposits of Devensian date have been investigated elsewhere in the valley of the River Great Ouse, generally providing evidence for open, tundra-like cold climates. Plant macrofossil assemblages from Late Devensian sands and gravels at Block Fen (West et al. 1995) provided similar evidence for open landscapes, with the most abundant taxa coming from environments interpreted as open, dry-ground herbs, with aquatic taxa also frequently recorded, including several species of pondweed. Betula (birch) and Salix (willow) were also recorded, although tree and shrub taxa were rare from these deposits. Similarly, plant macrofossil assemblages from Middle Devensian deposits at Somersham (West et al. 1999) have provided evidence for full glacial conditions, with a rich assemblage of dry, open-ground, damp-ground and occasional aquatic taxa. Tree taxa were rare, but again included birch and willow, with common dogwood recorded as part of the shrub taxa.
Richly organic deposits from the River Ouse (a tributary of the Great Ouse) at Earith, Huntingdonshire (Bell 1970), provided evidence of pools in a floodplain landscape, radiocarbon dated to 42 ky. Here, a similar assemblage of dwarf willows and herbs of base-rich, open habitats was recorded, with a mixture of northern and southern distributional types, the latter including species requiring a July mean temperature approaching 16℃, although mean annual temperatures were interpreted to be slightly below 0℃ (Bell 1970). Organic muds, considered likely to be of mid-Devensian date, were recorded as a channel fill in the valley of the Great Ouse at Radwell, Bedfordshire (Rogerson et al. 1992). Plant macrofossil and other biological remains in these organic deposits were interpreted as forming in interstadial conditions, although this reconstruction was complicated by the fluvial nature of the sediments (Rogerson et al, 1992).
Seven samples from waterlogged deposits forming part of Association B were scanned for insect remains, with the primary purpose of identifying beetle (Coleoptera) species that could be used for climate reconstruction using the mutual climatic range (MCR; Atkinson et al. 1987) system, as implemented in the BugsCEP program (Buckland and Buckland 2006; Buckland 2007). Five samples were from floodplain deposits in section SS05 described as root beds (samples 780095, 780096, 780058, 780099) and organic mud sample 780094, and one was from a deposit sampled primarily for clast lithology, sample 780132. OSL dates from interbedded sand and overlaying deposits in section SS05 indicated that these deposits spanned the period 45–40 ky, but with substantial overlaps in standard errors and some date inversions. A final sample, 780014 came from a disturbed/reworked root bed in section SS02 that also formed part of Association B. AMS results for this sample indicated a date of >38.9 ky (View report in digital archive [PDF]).
The samples were prepared using standard techniques (e.g. Kenward et al. 1980) and scanned in industrial methylated spirits (IMS) in a Petri dish using a stereoscopic zoom microscope (×7–×45). Where necessary to identify species that could potentially provide MCR data, selected beetle sclerites were picked out onto moist filter paper for close examination (chiefly Carabidae, Staphylinidae and Aphodiinae). Identification was by comparison with modern reference material and by reference to standard published works (Luff 2007; Lane et al. 2020). The abundance of insect taxa was recorded semi-quantitatively. Nomenclature for Coleoptera follows Duff (2018).
Insect remains, predominantly beetles, were recovered in moderate to good numbers from all seven samples. Fragmentation of the remains was generally high, however, especially among the larger taxa. Notably, many larger ground beetles (Carabidae) and scarabaeid dung beetles (Aphodiinae) were represented by very fragmentary material. The material was generally very fragile, and a proportion of the remains in some samples showed significant signs of chemical erosion, resulting in thinning, changes in surface detail and colour loss. The assemblages were broadly similar in species composition and are discussed together below. Interpretation of local environmental conditions was limited because the assemblages were recorded by scanning and many taxa were not closely identified. The results of the scanning are presented in Table 4 (PDF).
Overall, the insect assemblage indicated an area with standing water, sparsely vegetated damp ground where vegetation included sedges (Carex), rushes (Juncus) and moss, and drier sandy ground. The wider floodplain appears to have been open grassland, and the abundance and diversity of the dung fauna strongly indicates significant populations of grazing herbivores.
The water beetle taxa that were identified closely are typical of still to slowly flowing water (Helophorus grandis, Agabus bipustulatus, Colymbetes cf. fuscus, Hydrobius fuscipes s.l.). Small Helophorus species were relatively common in most samples but were not closely identified. No obligate running water taxa were noted. Larval sclerites of caddis flies (Trichoptera), predominantly of one taxon, were common in some samples, and chironomid larval head capsules were present in three samples.
The wider range of taxa representing damp ground with sandy or clayish soils and waterside habitats included eight species of Bembidion. Bembidion quadrimaculatum, B. bruxellense, B. ?obliquum and B. bipunctatum are typical of sparsely vegetated damp places. Platystethus nodifrons and P. nitens typically live on water margins and in marshes with fluctuating water levels (Lott 2009, 42). Moss and plant litter are suggested by Bembidion ?biguttatum, Arpedium quadrum, Simplocaria and Byrrhus. Kateretes is found on sedges (Carex) and rushes (Juncus), and Donacia or Plateumaris, represented only by small fragments of cuticle in one sample, are associated with a limited range of aquatic and marginal vegetation.
Notiophilus aquaticus, one of the commoner ground beetles (present in five samples), is typical of poor, sandy, dry or disturbed soils, often near water (Luff 1998, 25). Atomaria species, common in one sample from an organic mud deposit sample 780094, are generally associated with relatively dry plant litter, for example in tussocks of vegetation, and may have washed into the mud from a drier area. A record of Saprinus immundus, an exclusively maritime species of clown beetle (Histeridae), is notable as it fits with pollen evidence for halophytic plants typical of coastal settings, including thrift (Armeria maritima) and sea plantain (Plantago maritima). The beetle occurs in open dune systems on carrion and dung. There are very few modern records but historically it was found on the East Anglian and Kent coasts, with a few old records from the coast of Lancashire (Lane 2020, 183–184). Possibly associated, Bembidion varium occurs on damp soils near water, most frequently (but not always) in salt marshes (Luff 1998, 77; Luff 2007, 84), and B. bipunctatum is also halotolerant. Saline conditions may have developed locally in a continental climate because of high evaporation rates (West 2000, 168).
There were no clear insect indicators for either dead wood habitats or trees and shrubs among the identified taxa, although tiny decayed leaf fragments were noted in the paraffin flot from sample 780132. The remaining beetle taxa chiefly indicated a generally open environment, grassland habitats and populations of grazing mammals. Substantial numbers of scarabaeid dung beetles (Aphodiinae spp.) and other beetles primarily associated with dung (e.g. Cercyon melanocephalus, Cryptopleurum minutum, Anotylus gibbulus group, Oxytelus laqueatus or piceus) accounted for a major part of the terrestrial beetle fauna in each sample. The aphodiine dung beetles included Acrossus rufipes, two Chilothorax species and Melinopterus prodromus, but other unidentified species likely included some not currently extant in the British Isles. All these beetles are usually dependent on the dung of large mammalian herbivores, but Melinopterus prodromus, one of the commoner species in some samples, will also exploit foul decaying vegetable matter or overwinter in flood refuse (Jessop 1986, 22). Pasture and dung habitats were also suggested by Platystethus arenarius, Tachinus rufipes and T. corticinus. Phytophages commonly occurring in grassy habitats included Sitona species, represented in all the samples and associated with leguminous plants (Fabaceae), and Longitarsus species, and, especially associated with members of the Boraginaceae, Convolvulaceae, Plantaginaceae, Scrophulariaceae, Laminaceae and Asteraceae (Duff 2016, 156), Galeruca tanaceti, and Graptus triguttatus. The last species most typically occurs on ribwort plantain (Plantago lanceolata). Aclypea undata, a phytophage found at the bases of plants and now considered extinct in Britain, is known from open habitats on the European continent, including pasture grassland (Lane et al. 2020, 217–218).
The MCR method (Atkinson et al. 1987) uses thermal range data from predatory or scavenging species of beetles. Phytophages are not used because they are dependent on the distribution of their host plants. The method reconstructs the thermal environment in which the largest proportion of species represented in a sample could survive. Mean air temperatures of the warmest month (Tmax) and coldest month (Tmin), and the difference between Tmax and Tmin (Trange), can be calculated, which can indicate the degree of continentality. A range of temperatures is produced for Tmax and Tmin, rather than average temperatures such as can be calculated from modern data based on actual climatic records.
Climatic tolerance data exists for 26 species identified from the current site, chiefly Carabidae and Staphylinidae, and mean monthly winter temperatures (Tmin) of –11° to 4℃, and summer temperatures of 16–17℃, were estimated. Modern means for the Huntingdon–Cambridge region are 4.5–4.6℃ in January and 17.4–17.6℃ in July (Climate-Data.org 2023;). The MCR data obtained from the ancient assemblages suggests that summer warmth was well within the range of present-day temperatures. Winter temperatures were probably somewhat cooler, with consequent increased periods of frosts and snow cover. This fits with evidence of colder conditions suggested by the pollen work (Section 7 above) with the caveat that the available dataset for the beetle fauna was produced by scanning, and some taxa were too poorly preserved for identification to species level. The greater difference between maximum and minimum temperatures indicated for the current site suggests a more continental climate than currently.
Indications of the prevailing climate from the individual species among the identified beetle fauna are somewhat equivocal. The ground beetles Notiophilus aquaticus and Bembidion bipunctatum, both represented in several samples, are today generally commoner in northern parts of Britain, perhaps suggesting relatively cool temperatures, but conversely B. gilvipes, also well represented, is currently regarded as a southern species (Luff 1998, 25, 53, 59). Arpedium quadrum, a predatory omaliine rove beetle found among moss and plant debris (Zanetti et al. 2016), is no longer present in Britain but there are records from Quaternary deposits prior to MIS 5e (>c. 130 ky), late glacial (c. 15–10 ky) and Holocene deposits (Buckland and Buckland 2018). Modern records are from more northerly parts of Europe, especially Scandinavia and Finland, although some extend as far south as the extreme north of Italy (some extend as far south as the extreme north of Italy). The Anotylus gibbulus group, recorded from five samples, appears to be associated with dung of large mammals. It is widely recorded from Pleistocene deposits in Europe (and also from Toronto, Canada), but a major contraction in range appears to have occurred during the Devensian period and it now seems to be restricted to the Caucasus mountains and eastern Siberia north of Vladivostok (Hammond et al. 1979; Elias 1994, 66). As fossils the species has been particularly recorded from MIS 7 interglacial deposits, where it is often the most abundant rove beetle (Staphylinidae), e.g. Stanton Harcourt, Oxfordshire (Briggs et al. 1985) and Marsworth, Buckinghamshire (Green et al. 1984). There are also records from later deposits, including from interstadials within the Devensian glaciation. The species appears to have an association with the dung of large herbivores, and it has been suggested that its drastic contraction in range since the end of the Pleistocene may be in response to the extinction of many large mammal species (Morgan and Morgan 1980).
A Chilothorax species (an aphodiine dung beetle) with a distinctive pattern of elytral maculae was recorded from at least three samples. The remains were closely similar to both C. distinctus, found in present-day Britain, and C. jacobsoni, which currently occurs in Mongolia and southern Siberia (Frolov 2001; Akmetova and Frolov 2014). The elytral pattern is somewhat variable within both species. Chilothorax jacobsoni has been recorded from other deposits dated to MIS 3 on the Thames terraces in Oxfordshire and at Finningley in South Yorkshire (Briggs et al. 1985; Buckland et al. 2019), where it is suggestive of cool conditions, and possibly from Sandy in Bedfordshire (Gao et al. 1998). There is no thermal tolerance data for either species but C. jacobsoni would be expected to be indicative of colder conditions based on its modern range.
It was not possible to use the MCR method for the beetle assemblage from Lynford, Norfolk, but estimates based on modern geographical ranges of the species represented suggested that summer temperatures may have been around 12–14℃, with winter temperatures perhaps in the region of –8 to –15℃ (Coope 2012). Buckland and Buckland (2018) emphasizing that the overall beetle data set for MIS 3 (c. 59–28 ky) in England indicates substantial variations in summer and winter climates, which is consistent with evidence obtained from the Greenland ice-core record (Wolff et al. 2010; Rasmussen et al. 2014). They also stress that correlations between sites is problematic because of the imprecision and uncertainties involved with dating terrestrial sites of these periods.
Molluscs were recorded in low quantities in samples from organic and palaeosol deposits in sections SS05 and WkS06. Shells typically comprised small broken and unidentifiable fragments of indeterminate taxa with little or no palaeoenvironmental potential for further analysis or study.
A total of 120 faunal specimens was examined for this report (View report in digital archive [PDF]). The specimens comprised an assemblage derived from in situ Association A (n = 50 specimens) and Association B (n = 14 specimens) deposits and reject pile contexts (n = 28 specimens) within the north cell extraction area (Figure 1). A further 28 specimens were also recovered from the south cell area. From these, 96 were identified to species, genus or at least family, while 24 specimens remained unidentified.
This report presents an overview of the TEA28 assemblage in terms of taxonomical composition and taphonomic history, as well as some relevant details on the remains of each taxon in terms of skeletal element representation, ageing and sexing, pathology, and osteometry.
Five species were represented, all large or very large taxa: woolly mammoth (Mammuthus primigenius), woolly rhinoceros (Coelodonta antiquitatis), bison (Bison priscus), horse (Equus ferus) and reindeer (Rangifer tarandus) (Table 5).
| Taxon | No. of specimens |
|---|---|
| Mammuthus primigenius (Blumenbach 1803), woolly mammoth | 53 |
| Equus ferus Boddaert, 1785, horse | 3 |
| Coelodonta antiquitatis Blumenbach (1807), woolly rhinoceros | 17 |
| Rangifer tarandus Linne, 1758, reindeer | 2 |
| Cervidae indet. | 1 |
| Bison priscus Bojanus, 1827, steppe bison | 8 |
| Large bovid (Bos primigenius/Bison priscu, aurochs/steppe bison) | 10 |
| Cervidae/Bovidae | 2 |
| Mammalia, indet. | 24 |
The absence of medium and small mammals is, likely, a result of their removal by flowing water. Voorhies' experiments (Voorhies 1969) studying the behaviour of bones as sedimentary particles in flowing water show that different elements require different minimum current velocities to be transported, according to their size, density and shape (Behrensmeyer 1975). Voorhies grouped elements that tend to be transported together into three categories, groups I, II and III, with the velocity required to transport elements increasing from the former to the latter. An assemblage containing all three groups can be regarded as relatively undisturbed; assemblages containing groups II and III can be regarded as 'winnowed'; if elements belonging to group III predominate, they represent a 'lag' deposit. However, it must be stressed that Voorhies' experiments were carried out with moderately large vertebrates; bones of larger mammals such as rhinoceros and elephant require much higher flow velocities than the analogous skeletal elements of smaller species.
The TEA28 assemblage included both cranial and postcranial material (Table 6). Among the latter, some elements are notoriously absent/underrepresented (relative to their abundance in the skeleton): phalanges, carpals and tarsals, vertebrae and, to a certain extent, ribs. These are elements found in groups I/I–II, i.e. they are among those most susceptible to transportation by fluvial action (Voorhies 1969). The most common elements represented in the assemblage belonged nominally to Voorhies' groups II and III. However, given that most specimens nominally in group II (e.g. femur, humerus) are in fact from winnowed groups, it must be noted that, while TEA28 can be regarded as a basically non-transported assemblage, approximately a quarter of the specimens showed signs of abrasion, for example rounding or polishing of the bones and exposure of trabecular tissue, which may be a useful characteristic for recognising bones that have been transported in fluviatile environments. However, abrasion can also be caused by sediment movement around stationary bone (Behrensmeyer 1982), which is the interpretation preferred here.
| Taxon | Mammuthus | Coelodonta | Bison and Bos/Bison | Equus | Rangifer + Cervid | Bovid/Cervid |
|---|---|---|---|---|---|---|
| Antler | _ | _ | _ | _ | 2 | - |
| Skull | 2 | 6 | 1 | - | - | - |
| Horn core | _ | _ | 1 | _ | _ | _ |
| Tooth (tusk) | 27 | _ | _ | _ | _ | _ |
| Tooth (other) | 3 | - | - | - | - | - |
| Mandible | 2 | - | - | - | - | - |
| Scapula | 1 | 1 | 1 | - | - | - |
| Humerus | 3 | 1 | 5 | - | - | - |
| Radius | - | 1 | 1 | 1 | - | - |
| Ulna | - | 2 | - | - | - | - |
| Pelvis | 1 | 3 | - | 1 | - | 1 |
| Femur | 2 | 1 | - | - | - | - |
| Tibia | 1 | 1 | 5 | 1 | 1 | 1 |
| Fibula | 1 | - | _ | - | _ | _ |
| Calcaneus | 1 | - | 1 | - | - | - |
| Metatarsal | - | - | 2 | - | - | - |
| Metapodial | - | - | 1 | - | - | - |
| Vertebra | 1 | - | - | - | - | - |
| Rib* | 2 | 2 | - | - | - | - |
| Unidentified element + | 6 | - | - | - | - | - |
| Total | 53 | 17 | 18 | 3 | 3 | 2 |
As the assemblage appeared to represent basically undispersed remains, it retained more palaeo-ecologic information than assemblages composed primarily of only group I or only group II elements (Behrensmeyer 1975). The presence of cold or cool-adapted taxa, i.e. the woolly mammoth, horse, woolly rhinoceros, reindeer and bison, in TEA28 is consistent with the assemblage falling within Currant and Jacobi's Pin Hole mammal assemblage zone from the mid-Devensian, c. 60–27 ky Currant and Jacobi 2011). The association of woolly mammoth, horse, woolly rhinoceros, reindeer and bison is thought to indicate a landscape with abundant grasslands and very few trees (Scott 2018), this being particularly true for Coelodonta, which was a specialised grazer in open grasslands, as indicated by morphology, teeth mesowear and 13C values (Pushkina et al. 2020). Pollen and plant macrofossils seem to confirm a very open environment for TEA28. It must be emphasised, however, that fluvial bone assemblages typically represent time intervals in the order of 102–104 years, so the collection retrieved from TEA28 does not represent a 'living community' but rather a thanatocoenosis, i.e. animals that came together at death, or, even more likely, a taphocoenosis, i.e. animals that came together after death.
While mostly non-transported, characteristics in the assemblage still point towards a complex taphonomic history, attested for example through variable weathering between specimens. There was no apparent correlation between their location (association, context) and their degree of weathering. Bone material in fluvial channel deposits is commonly derived from multiple sources: 1) autochthonous osseous material in the floodplain soil that is incorporated by partial destruction and/or reworking through bank erosion; 2) overland transport directly into the channel; and 3) carcasses or bones originally present in the channel (Behrensmeyer 1982).
There is possibly an example of the latter at TEA28: a femur, tibia, fibula and calcaneum of Mammuthus, all with unfused epiphyses, and all from the left hand-side, were recovered from basal gravels. All but the calcaneum came from a single context. These specimens must be seen as parts of the hindlimb of one immature mammoth. This suggests that the leg was deposited in the fluvial channel while still in articulation, or, at the very least, the bones remained in close association (e.g. on the floodplain) until they were reworked into the channel. In any case, this indicates that the mammoth died not far from the find location.
There is some evidence to suggest that large mammals frequented the site: ten specimens presented multiple, shallow linear marks usually oriented in a transverse direction to the long axis of bones. This type of mark has been observed on elephant bones in areas where elephant traffic is frequent (Haynes et al. 2020). These marks are thought to be a result of the trampling of bones resting on a stony substrate by elephants and other large mammals, and this is likely to be the case for TEA28 (Schreve 2006; Fernandez-Jalvo and Andrews 2016; Haynes et al. 2020).
No signs of hominin agency in the form of butchery marks were observed in the assemblage, but carnivore gnawing was visible on two specimens: on the ventral end of a mammoth rib, and on the caput of a woolly rhinoceros femur. This indicates that at least some of the remains were exposed and available to scavengers for a period of time before being buried.
Of the 53 mammoth specimens, just over half were tusk fragments. They were invariably weathered, although the degree of weathering varied. It was not possible to assign an approximate age to any of the tusk remains but, given that tusk diameter appears to be strongly sexually dimorphic in woolly mammoth (Lister 2009), the sex of some of the specimens could be assigned with a degree of certainty. Of the 14 measurable tusk specimens, three very large specimens could be assigned as males, whereas the remaining, smaller tusks may have belonged to adult females or immature males.
A fragment of the tip of a tusk, still preserving some of the outer layer of cementum, had a smooth, polished surface on an old, fractured surface, known as 'faceting' (Figure 21). In living proboscideans, broken tusks sometimes occur during, for example, fights between males, or when elephants of both sexes shove each other to reach critical resources such as water. The fracture surface of the remaining (rooted) tooth then becomes smoothed from use (Haynes et al. 2020). It is very likely that this also occurred in extinct proboscideans such as mammoths (Haynes 2020).

Two essentially complete teeth were recovered, as well as an additional fragment of a molar or deciduous premolar. An upper right third molar represented an individual older than about 40 years, whereas an upper left first molar could be assigned to an individual just under 10 years of age. The younger individual probably had not yet attained sexual maturity, which does not happen in living elephant taxa, and thus probably in mammoths, until the age of 12–15 years (Haynes 1991).
Among the postcranial elements found in the basal gravels, some were part of the same left hindlimb of an immature individual. The size of the specimens suggests that they did not belong to a very young animal. Based on the age of epiphysial fusion in modern African elephants (Loxodonta africana), the remains would represent an individual somewhat younger than c. 20 years, if it was a female, but somewhat younger than 28–30 years if it was a male (Haynes 1991; Lister 1999). The fact that these remains were in association strongly suggests that the leg was initially deposited, probably in articulation, soon after the death of the individual, and that no significant transport affected these elements (it was impossible to establish whether the much smaller, missing epiphyses of the tibia and calcaneum were carried downstream by flowing water, or whether they were missed during the excavations).
Other postcranial finds of mammoth notably included a broken but nearly complete humerus with both epiphyses fused. Comparisons with male mammoths from Condover (Shropshire, UK) and Siegsdorf (Germany) (Lister 2009; Larramendi 2015), suggest that the Fenstanton specimens represent a skeletally mature female, at least 20–25 years of age.
Only three horse specimens were recovered. A radius and a tibia represented skeletally mature individuals, i.e. older than c. 4 years. A fragmentary innominate bone had a fused acetabulum. The radius and the pelvis showed somewhat rounded edges, possibly an indication that they had been transported by flowing water for some distance (but see above). Only two measurable specimens were recovered from Fenstanton, but their size relative to other British horses from MIS 3–2 is very difficult to assess, as no measurements have been published for this time span.
Fifteen specimens were assigned to this taxon, and a further two (ribs) were identified as probably belonging to this species. The specimens comprised both cranial and postcranial elements (Table 6, 2). The remains were derived from a minimum of six individuals, based on cranial elements (Figure 22).
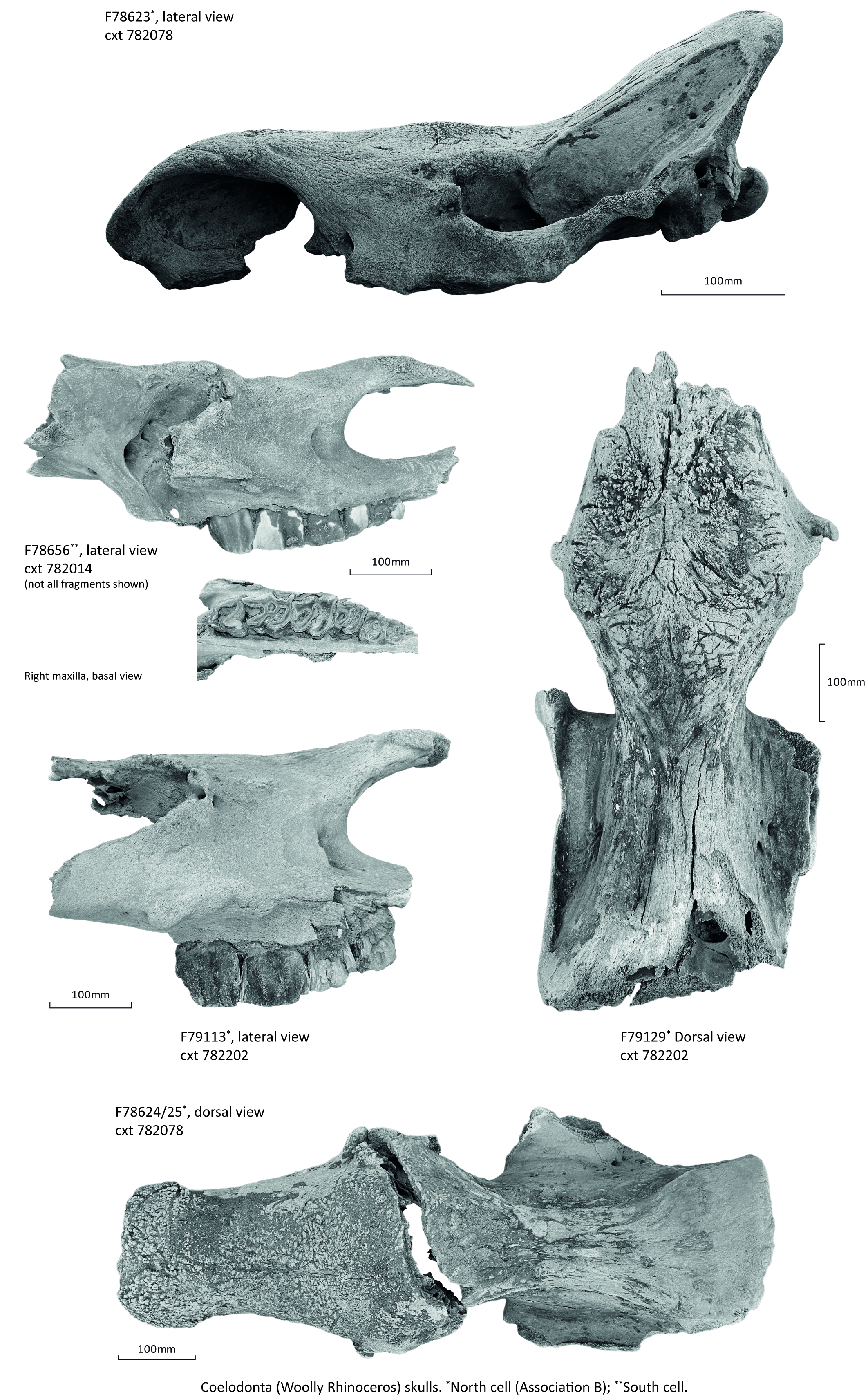
Two relatively well-preserved skulls, though lacking teeth, could be assigned, based on their nasal bones and septum, to older adults; a further two rather more fragmentary skulls, with some teeth present, could be assigned to somewhat younger individuals, probably 14–20 years of age, based on analogous tooth wear in the African white rhinoceros (Hillman-Smith et al. 1986).
The presence of both older and younger adults is also suggested by the postcranial material. Osteometrically, Coelodonta at this site seemed to fall consistently around the lower end of the size range recorded in Eurasia for this species. It is still unclear whether this indicates that the woolly rhinoceros in Britain was smaller than its Eurasian counterparts; this can only be clarified with additional osteometrical data from British specimens.
Eight specimens were assigned to the steppe bison rather than aurochs (Bos primigenius), based on published morphological criteria. A further 10 were assigned to 'Bos or Bison', though it is unlikely they represent the former, as 1) no unambiguous remains of aurochs were identified, and 2) this taxon seems to be absent from MIS 3 and 2 in Britain, although it is present during the Allerød Interstadial.
Based on the side of the body, size and epiphysial fusion, a minimum of three individuals was represented in the assemblage, but this number is very likely an underestimation. Most specimens seemed to represent mature animals, but an individual younger than c. 42 months was also present. Metrical data was obtained from some of the specimens. Given that bovines, and especially B. priscus, show a large degree of sexual size dimorphism, such data allows probable sex determinations. Five specimens were assigned as males, four as females, and two as possible males.
Notable specimens of this taxon included the frontal bone of a skull with a large fragment of the right horn core (Figure 23). While the horn core was incomplete and thus its length could not be measured, its basal oro-aboral diameter corresponded to horn cores of male B. priscus found at the site of Habarra, in the French Pyrenees, dating to MIS 3–2, suggesting that the specimen from Fenstanton was a bull. Interestingly, the horn core showed an elongated focal resorptive. Similar lesions have been observed in domestic cattle and, more significantly, in a Pleistocene (MIS 3) steppe bison from Germany (Kierdorf et al. 2016). The aetiology of this pathology is unclear; potential causes include local pressure-induced bone resorption (e.g. as a result of trauma) or a systemic origin, for example caused by periods of nutritional (calcium, phosphorous) deficiency.
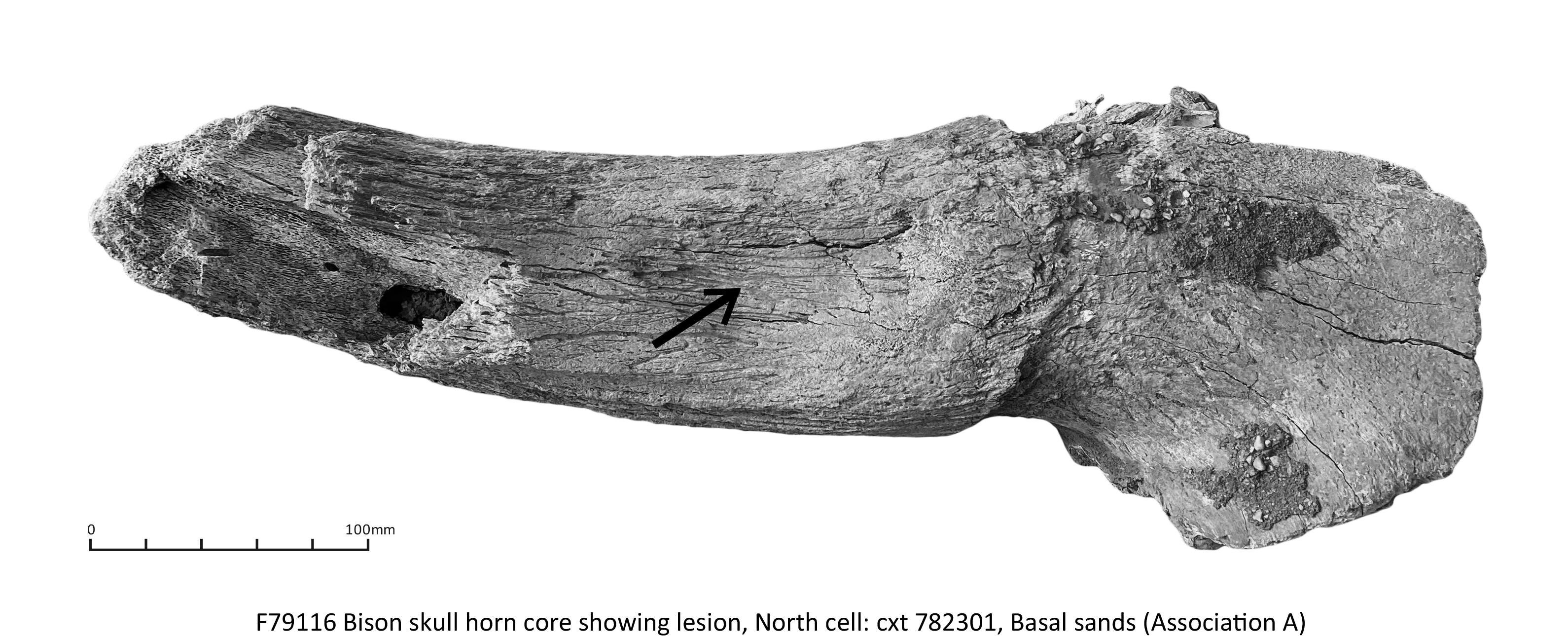
Only two specimens of this taxon were recovered, both shed antler fragments. They comprised the burr and the proximal part of the main beam. One of the specimens showed some rounding of its surfaces, indicating a degree of transportation by water. One of the specimens was relatively large and could represent a male, whereas the other was somewhat smaller and therefore likely to represent an adult female or a subadult individual.
The faunal assemblage from TEA28 is taphocoenose, representing mostly lag deposits. It includes cold/cool-adapted taxa: mammoth, woolly rhinoceros, bison, horse and reindeer. This is consistent with Currant and Jacobi's Pin Hole mammal assemblage zone from the mid-Devensian, c. 60–27 ky. No human agency was observed on any of the specimens, whereas carnivore gnawing could be seen on a small number of specimens; this, together with light to moderate weathering on most specimens, indicates that the material was not buried very rapidly, but was exposed instead to the elements and scavengers.
Single finds of lithic artefacts were recovered from a small number of deposits in both the north and south cell areas of the borrow pit. On-site, individual finds were assigned object and deposit context numbers, with coordinate and elevation (OD) values recorded by GPS where practical and safe to do so. Off-site, artefacts were examined and information recorded for each regarding stratigraphic context, type, condition, edge damage, patination and staining. Where required, identifications were assisted by reference to illustrations in Wymer (1968), Tyldesley (1987), Debénath and Dibble (1994) and Bordes (2002).
Artefacts recovered by the watching brief comprised a small lithic assemblage of some seven pieces, from both in situ and reject pile contexts, with no primary context material identified. The assemblage included one complete and one broken handaxe (F79114 and F79132), one cleaver (F79174), a single bifacial scraper (F79115) and three unretouched flakes (F79131, F79133 and F79134). The complete handaxe, bifacial scraper and two unretouched flakes (F79131 and F79134) were recovered in situ from the gravelly sands and sandy gravels of facies Association A, and the handaxe fragment in situ from sandy floodplain deposits of Association B. One unretouched flake was also collected in situ from basal sand and gravel within the south cell extraction area (F79133). The cleaver was recovered from a reject pile in the north cell. The angular flint recorded for micromorph sample 780127 as a possible stone tool was a partially cortical natural flake with no evidence for human modification. Striking platform, bulb of percussion, retouch flaking scars and other modifications were not found on the piece.
All finds were made of flint, with raw material varying from light brown–grey banded flint to dark brown flint with grey–-light brown inclusions. Patination was present on the complete handaxe (F79114), with the upper face displaying a diffuse web-like grey–light grey patina and the lower face unpatinated, though with minor light brown–yellowish brown staining (iron oxide staining). This staining also occurred on five of the remaining artefacts, with three heavily stained (F79133, F79134 and F79174) and one flake displaying diffuse brown–yellowish brown staining (F79131). Both faces of the handaxe tip fragment (F79132) were also covered by mottled brown–yellowish brown staining. The bifacial scraper (79115) was unpatinated and not stained. Fluvial abrasion ranged from minor arris and flake scar edge rounding (F79114, F79115, F79174, F79134, F79131) to medium rolling with arris and flake scar edges displaying rounding, scratching and glossy surface polish (F79132, F79133). Recorded edge damage varied between moderate (F79114, F79115, F79174, F79131 and F79134) and severe (F79132 fragment, and F79133 platform missing), with edge-nicks and small flake removals present on edges (break surface, tip, both laterals and upper and lower faces) of all seven artefacts.
The handaxe was a typological variant of the bout coupé-type handaxe, with a slightly convex butt, dated to the late Middle Palaeolithic (Tyldesley 1987, 111, tables 3 and 4, 165). It was bifacially worked and symmetrical in planform, with a slightly convex butt edge and a rounded tip. Sides were slightly convex, with the intersection of the sides and butt formed by small shaping retouch flakes. Flat and narrow flake scars indicated soft hammer flake removal, with flake scar patterning indicating that it was made by working one face (upper) along one edge first and then by working the other edge; flake scars for the butt and tip showed that they were then worked. The other face (lower) appeared to have only been used for straightening or shaping the intersection between the sides and butt. The piece was possibly unfinished as 'retouch' on the lower face was largely limited to edge straightening flaking. Flake scars included large and small scalar and stepped; no tranchet scar was present.
This artefact appeared to be a small variant of the Quina-type bifacial scraper Bordes type 28: racloir à retouche biface type Quina (Bordes 2002, 47 figs 29, 1-2). It was made from a small side-struck flake with a cortical platform, and the bulb of percussion and bulbar scar removed by retouch/thinning. It was fully bifacial and asymmetrical in planform, with a rounded convex right lateral formed by small–medium shaping retouch flakes (30°edge angle) Flake scars included large and small scalar and stepped, with scalar largely on dorsal edges and stepped on ventral edges. Retouch was also present on the right ventral edge and dorsal distal edge; flake scars for these were typically small scalar and stepped, with use-ware and edge-damage scars indistinguishable.

The tip fragment was fully bifacial and slightly asymmetrical in planform, with a rounded tip and sides straight–slightly convex. Flake scars included large and small scalar and stepped; no tranchet scar was present. The characteristics of the related handaxe for the fragment remain unclear as only the tip was recovered and not the complete handaxe.

The cleaver was manufactured from a large flint thermal fracture with a cortical (84%) upper surface. Marginal bifacial retouch was present along the right lateral, with invasive retouch on the lower thermal fracture surface. Asymmetrical in planform, with slightly convex left lateral and straight-edged right lateral, and a rounded edge-damaged partial cortical butt, its distal cutting edge was unworked. Flake scars included large and small scalar. Typologically the artefact was classified as Wymer Type H: Cleaver and Bordes Type 55: Hachoir (Wymer 1968, 55–56; Bordes 2002, 85–88, figs 48, 71–78).
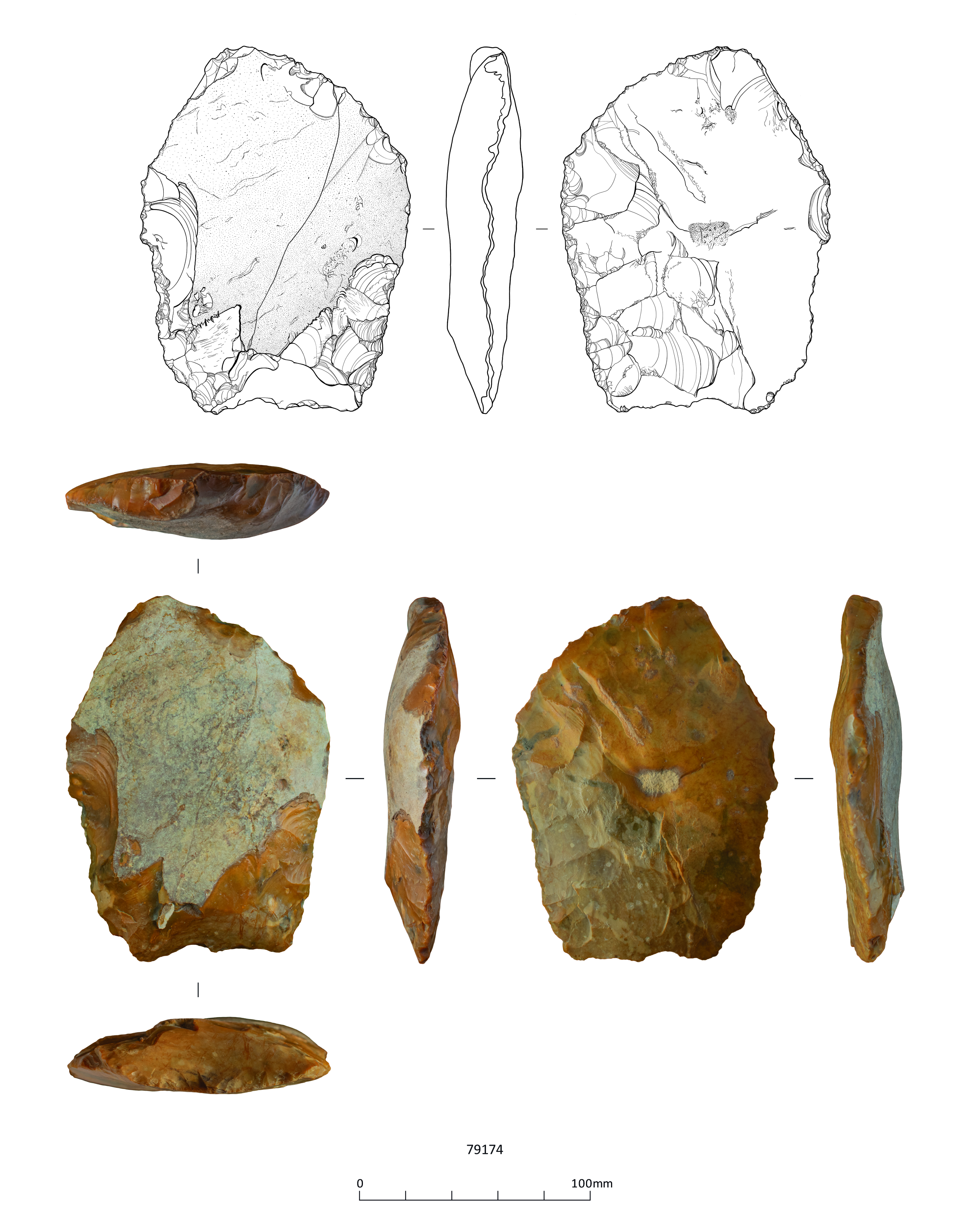
All three of these artefacts were unpatinated and exhibited brown–yellowish brown staining. Two of the flakes were non-cortical, with one having a flat striking platform, bulb of percussion and bulbar scar (F79131). The striking platform and most of the bulb of percussion were missing on the other non-cortical flake (F79133). The remaining unretouched flake (F79134) was partially cortical (45%), with part of the platform and bulb of percussion missing (proximal end with concave flake scar from spontaneous removal/shattering of the platform when struck). Dorsal flake scars ranged from two to four on individual flakes, with distal hinge, step and possible concave or angular step terminations also present. Use-ware and edge-damage scars were indistinguishable.
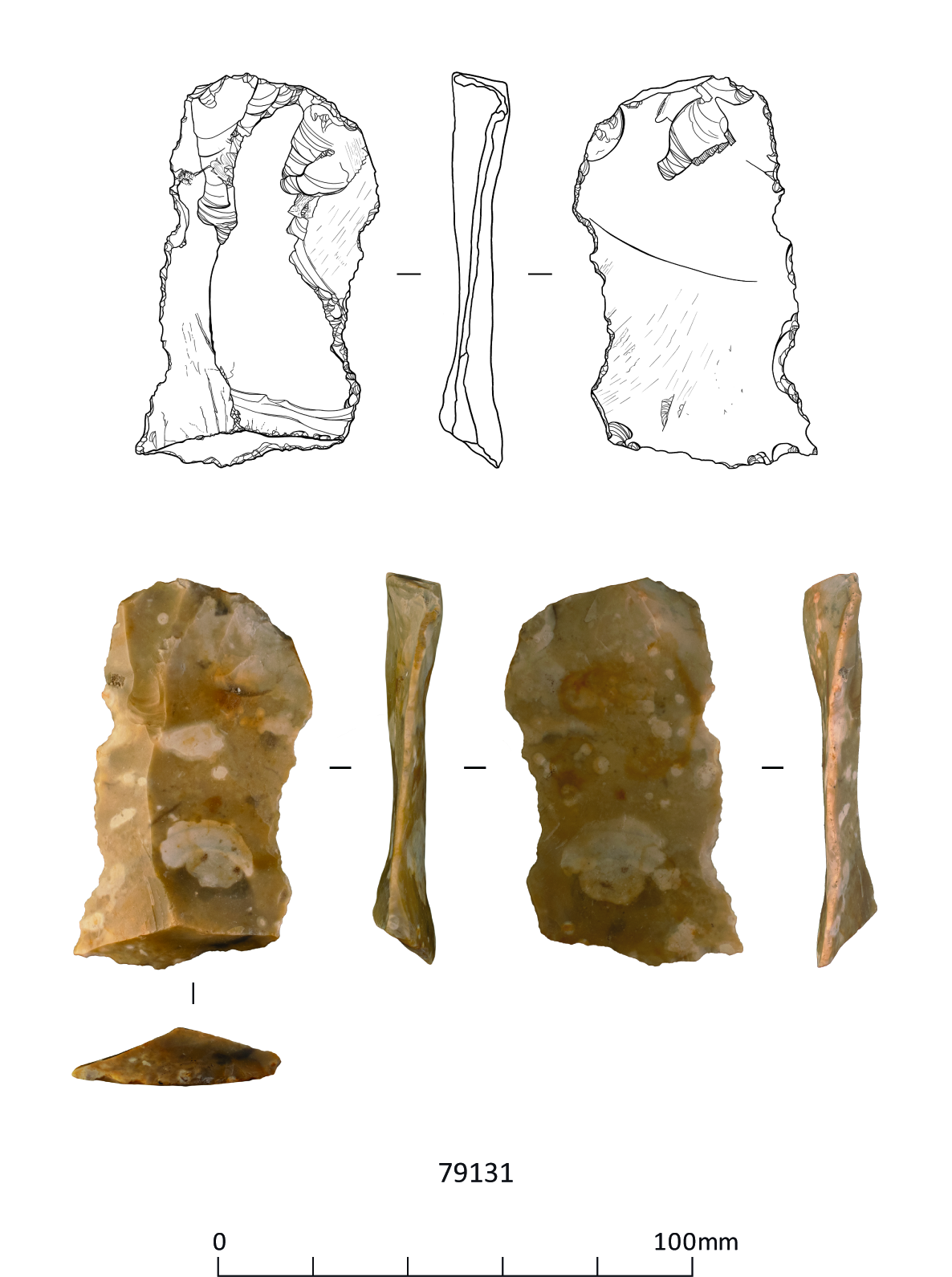
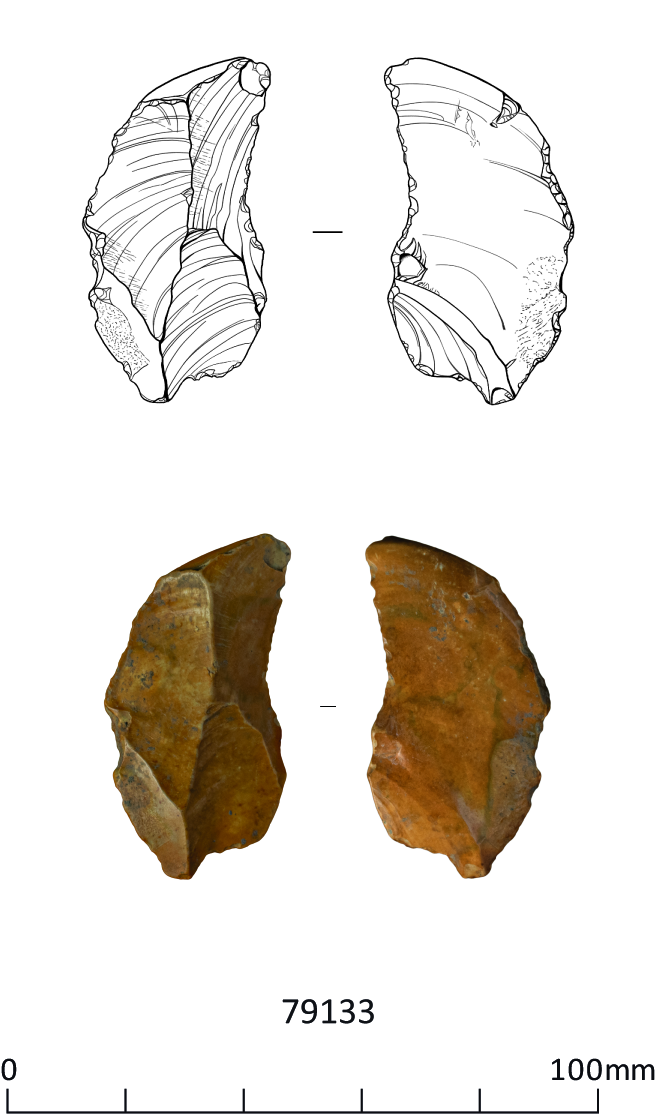
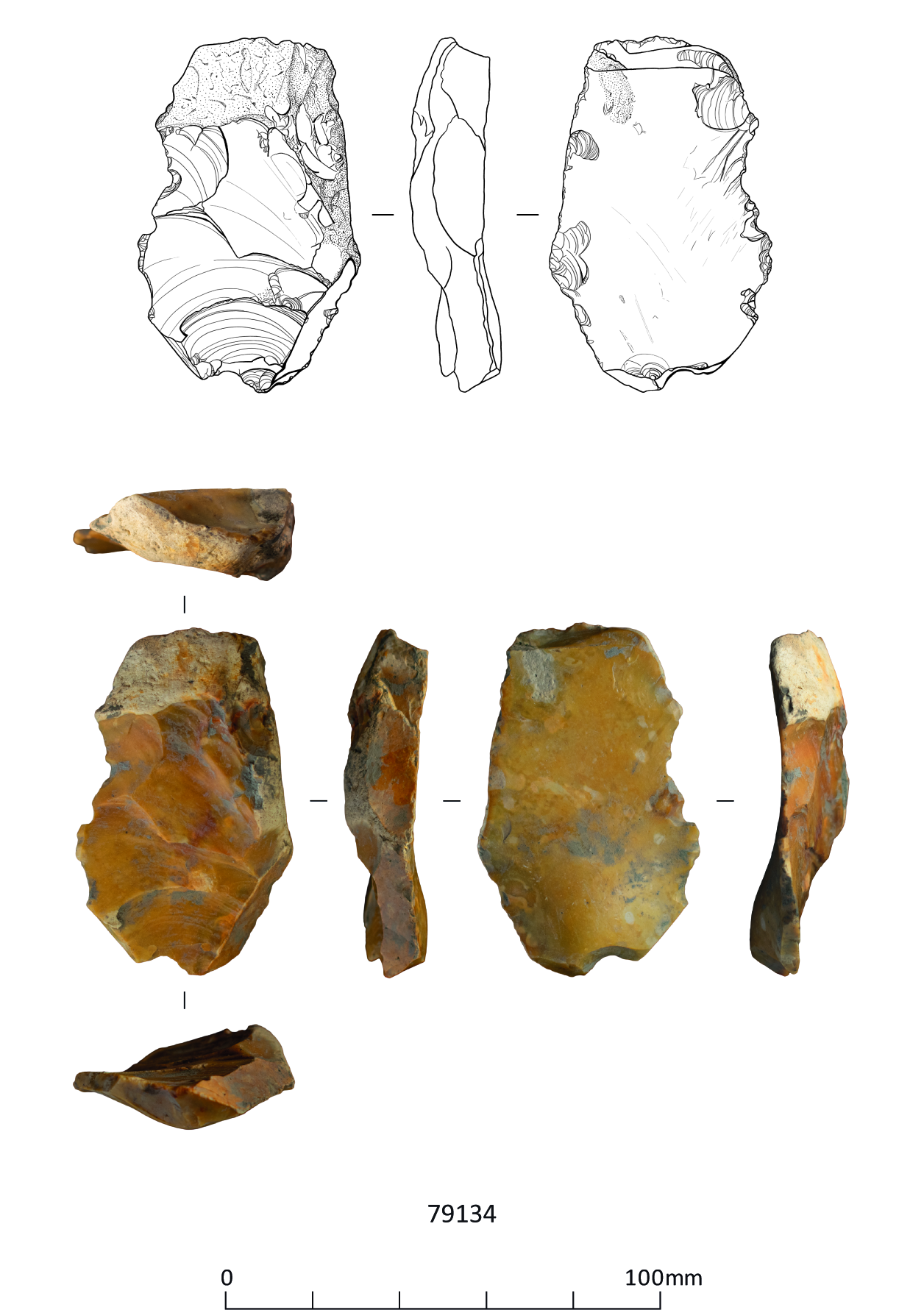
The Palaeolithic record for the extraction area comprised abraded and reworked lithic artefacts within younger river terrace deposits, with no in situ primary context archaeological material found for recorded sample sections. OSL age estimates of c. 43–38 ky for deposits with artefacts in them indicated that they dated to the Middle Devensian (MIS 3; c. 60–27 ky), falling within a transitional period of climate instability (c. 44–37 ky) identified on the GISP2 Greenland ice-core climate record lasting from the end of the British late Middle Palaeolithic period at c. 44 ky till the start of an early cold-stage at c. 37 ky (Van Andel et al. 2003; Boreham et al. 2010). The complete handaxe and bifacial scraper were typologically dated to the late Middle Palaeolithic, while the handaxe tip fragment was most likely Lower Palaeolithic in age (Tyldesley 1987; Bordes 2002). The small size of the lithic assemblage and its multiperiod make-up indicated that it represented an accumulation of reworked artefacts produced by fluvial erosion and sediment deposition processes, rather than any in situ human activity. Typological dates also suggested that the artefacts represented a time-averaged collection of material documenting a human presence in the region during the Lower and late Middle Palaeolithic.
A total of 21 samples was taken for luminescence dating from sections SS03, SS04 and SS05 in the north cell extraction area (Table 7). Fifteen samples from selected deposits were then chosen for dating to provide a chronological framework for the stratigraphic sequence of deposits. Samples were collected in opaque PVC tubes hammered into vertical sections, with in situ radioactivity measurements made by a gamma-ray spectrometer and calibrated against the Oxford blocks (Rhodes and Schwenninger 2007). Laboratory procedures were designed to produce sand-sized quartz (180–255 μm) and followed a standard preparation procedure (Schwenninger 2020). All laboratory treatments were carried out under low-intensity laboratory safe-lighting. Measurements were performed in an automated luminescence reader (model TL/OSL DA15 made by Risø; Bøtter-Jensen 1988, 1997, Bøtter-Jensen et al. 2000) using a single aliquot regenerative (SAR) post-IR (infrared) blue OSL measurement protocol (Murray and Wintle 2000; Banerjee et al. 2001; Wintle and Murray 2006).
| Field code | Lab. code | Sample no. | Context no. | Facies association | Section no. | Age |
|---|---|---|---|---|---|---|
| FEN19-01 | X7446 | 780025 | 782210 | A | SS05 (3) | 41295 ± 3155 |
| FEN19-02 | X7447 | 780026 | 782210 | A | SS05 (3) | 38810 ± 2440 |
| FEN19-03 | 780027 | 782212 | B | SS05 (3) | ||
| FEN19-04 | X7449 | 780028 | 782211 | B | SS05 (3) | 41810 ± 4400 |
| FEN9-05 | 780029 | 782213 | B | SS05(3) | ||
| FEN19-06 | X7451 | 780030 | 782213 | B | SS05(1) | 51285 ± 3760 |
| FEN19-07 | X7452 | 780031 | 782213 | B | SS05(1) | 45475 ± 5130 |
| FEN19-08 | X7453 | 780032 | 782206 | C | SS05(1) | 43045 ± 3855 |
| FEN19-09 | X7454 | 780033 | 782206 | C | SS05(1) | 45860 ± 3140 |
| FEN19-10 | 780034 | 782214 | C | SS05(1) | ||
| FEN19-11 | 780035 | 782214 | C | SS05(1) | ||
| FEN19-12 | X7457 | 780036 | 782203 | D | SS05(1) | 22570 ± 1570 |
| FEN19-13 | X7458 | 780037 | 782203 | D | SS05(1) | 6895 ± 540 |
| FEN19-14 | 780038 | 782215 | B | SS05(1) | ||
| FEN19-15 | X7460 | 780039 | 782215 | B | SS05(1) | 28950 ± 5325 |
| FEN19-16 | X7461 | 780040 | 782242 | B | SS05(1) | 53205 ± 4280 |
| FEN19-17 | X7462 | 780041 | 782229 | B | SS05(1) | 40435 ± 2845 |
| Sample 1 | X7463 | 780020 | 782302 | B | SS03 | 40900 ± 2900 |
| Sample 2 | X7464 | 780021 | 782202 | A | SS04 | 44800 ± 4310 |
| Sample 3 | X7465 | 780022 | 782201 | A | SS04 | 43495 ± 3635 |
| - | 780023 | 782203 | C | SS04 |
Age estimates were based on sand-size quartz, with palaeo-dose determinations made using a multi-grain SAR-dose protocol (Murray and Wintle 2000; Banerjee et al. 2001). For each of the 15 samples, gamma dose rates were based on in situ gamma-ray spectroscopy measurements, with beta dose rates derived from concentrations of radionuclides by elemental analysis using ICP-MS/AES performed by a specialist and accredited laboratory (Actlabs) in Canada (Schwenniger 2020). The final OSL age estimate included an additional 4% systematic error to account for uncertainties in source calibration and measurement reproducibility. Dose–rate calculations were obtained using the dose–rate conversion factors of Guérin et al. (2011) and calculated using DRAC software (version 1.2; Durcan et al. 2015). The measured moisture content of the samples was considered to be representative of the mean water content during the burial period. The contribution of cosmic radiation to the total dose rate was calculated as a function of latitude, altitude, burial depth and an average overburden density of 1.9 g/cm as provided by Prescott and Hutton (1994).
The age estimates obtained from the samples indicated that deposits in the north cell dated from the Middle–Late Devensian period (MIS 3–2; c. 60–15.4 ky). These age estimates were not all in stratigraphic order, with a number of age reversals and mixed dating results for individual facies associations. Calculated ages for samples X7451 and X7461 appeared substantially older than those from deposits stratigraphically equivalent or lying below them. Samples X7452, X7453, X7454 and X7464 also appeared somewhat older, and sample X7460 younger, than stratigraphically related deposits. This discrepancy between OSL dates strongly implied a degree of age overestimation, indicative of partial bleaching problems and/or mixed grain populations. Inadequate bleaching and the fluvial reworking of older sediments into younger deposits were suggested for the four age estimates of 53205±4280 (X7461), 51285±3760 (X7451), 45475±5130 (X7452) and 44800±4310 (X7464) (Wallinga 2002; Bateman et al. 2003; Rittenour 2008), and chemical weathering for the two age estimates of 43045±3855 (X7453) and 45860±3140 (X7454) (Jeong et al. 2007; Jones et al. 2020). Age underestimation was also indicated for the date of 28950±5325 (X7460) and was likely related to chemical weathering and/or partial bleaching. Other sources of OSL age estimation errors included factors such as calibration errors and dose–rate determinations (Smart and Frances 1991). As a result, the age estimates for these seven samples were not considered to be reliable and were not used in the stratigraphic analysis.
The quartz mineral grains extracted from the remaining eight samples presented excellent signal characteristics, including low thermal transfer, good recycling and negligible infrared signals indicative of feldspar contamination. The samples were also characterised by high sensitivity to laboratory-induced irradiation, and individual measurements on sized quartz multigrain single aliquots (diameter = 4 mm) produced excellent growth curves. The samples displayed remarkably low overdispersion values, ranging from 5 to 15%, thereby suggesting adequate resetting of the OSL signal at the time of deposition. Age estimates for these samples were considered to be reliable and in agreement with the stratigraphic succession of deposits. They ranged from 43495±3635 (X7465) at the base of the succession to 22570±1570 (X7457) at the top or 6895±540 (X7458) immediately below the topsoil. These estimates also included multiple dates for associations A (X7465, X7446, X7447), B (X74449, X7463, X7462) and D (X7457, X7458), with the dates of 43045±3855 (X7453) and 45860±3140 (X7454) for Association C considered unreliable on account of likely overestimation from chemical weathering.
Four samples were taken for AMS radiocarbon dating from organic sediments in sections SS02 and SS05, and from a palaeosol in section WkS06. Processed bulk samples were subsampled for plant macrofossils and prepared following standard laboratory procedures for AMS radiocarbon dating at the Oxford Radiocarbon Accelerator Unit (ORAU) (Brock et al. 2010). These included further subsampling, chemical pretreatment for the removal of contaminants and stable carbon isotope and AMS isotope measurements.
Dating results for the four samples are summarised in Table 8. All four produced infinite dates and lay beyond the reliable limits of the radiocarbon method. These dates, as a result, represent minimum estimates of the true age. The dates indicated that the likely true age of the sediments was probably in excess of 38–41 ky.
| OxA code | Sample no. | Context no. | Facies association | Section no. | Material (species) | ƍ13C | Date |
|---|---|---|---|---|---|---|---|
| OxA-43555 | 780014 | 782205 | B | SS02 | Seeds (Ranunculus) | -27.25 | > 38900 |
| OxA-43556 | 780058 | 782216 | B | SS05(1) | Seeds (Ranunculus)> | -26.74 | > 36300 |
| OxA-43557 | 780084 | 782238 | B | SS05(2) | Seeds (Linum) | -25.31 | > 40600 |
| OxA-43558 | 780132 | 782403 | B | WkS06 | Seeds (Linum & luzula) | -27.15 | > 38800 |
The fluvial, floodplain and periglacial colluvial deposits recorded for the section samples form part of the Ouse Valley Formation (Green et al. 1999, 44; Boreham et al. 2010; Gao and Boreham 2011). OSL age estimates indicate that these deposits span the Middle–Late Devensian period (MIS 3–2; c. 60–15.4 ky), with facies Associations A, B and C being part of the Hemingford Member and dating to the Middle Devensian, at c. 43495±3635 (X7465) and c. 38810±2440 (X7447), and Association D being part of the St Ives Member, dating to c. 22570±1570 (X7457) and the last glacial maximum (MIS 2; c. 27–16 ky). Temperate deposits similar to those at nearby Woolpack Farm (Gao et al. 2000) and Galley Hill (Cambridge 1964, 1965; Forbes and Cambridge 1966; Preece and Ventris 1983), and dated to the Ipswichian Interglacial (MIS 5e; c. 127–115 ky), do not occur beneath river terrace sands and gravels within the borrow pit area.
The geometry of the channel is poorly known, with quarry sections suggesting that a shallow channel was incised into the Oxford Clay bedrock, and OSL dates suggesting that this incision may have occurred around c. 43.5 ky. Comparison with the oxygen isotope climate record from the GISP2 Greenland ice core shows that this date falls within a transitional period of climate instability (c. 44–37 ky) sandwiched between a stable warm phase (c. 59–44 ky) that is equated with the Upton Warren Interstadial, and an early cold phase (c. 37–27 ky) lasting to the start of the last glacial maximum (Figure 31) (Van Andel et al. 2003; Boreham et al. 2010; Boismier and Green 2012). This date suggests that channel incision may have occurred at the transition (cooling) between the Middle Devensian warm and cold phases, and later than the widespread fluvial downcutting at the MIS 4–3 transition noted for a number of other Fenland rivers (Boreham et al. 2010; Gao and Boreham 2011). Whether this channel incision was in addition to the MIS 4–3 downcutting or instead of it is currently poorly understood.
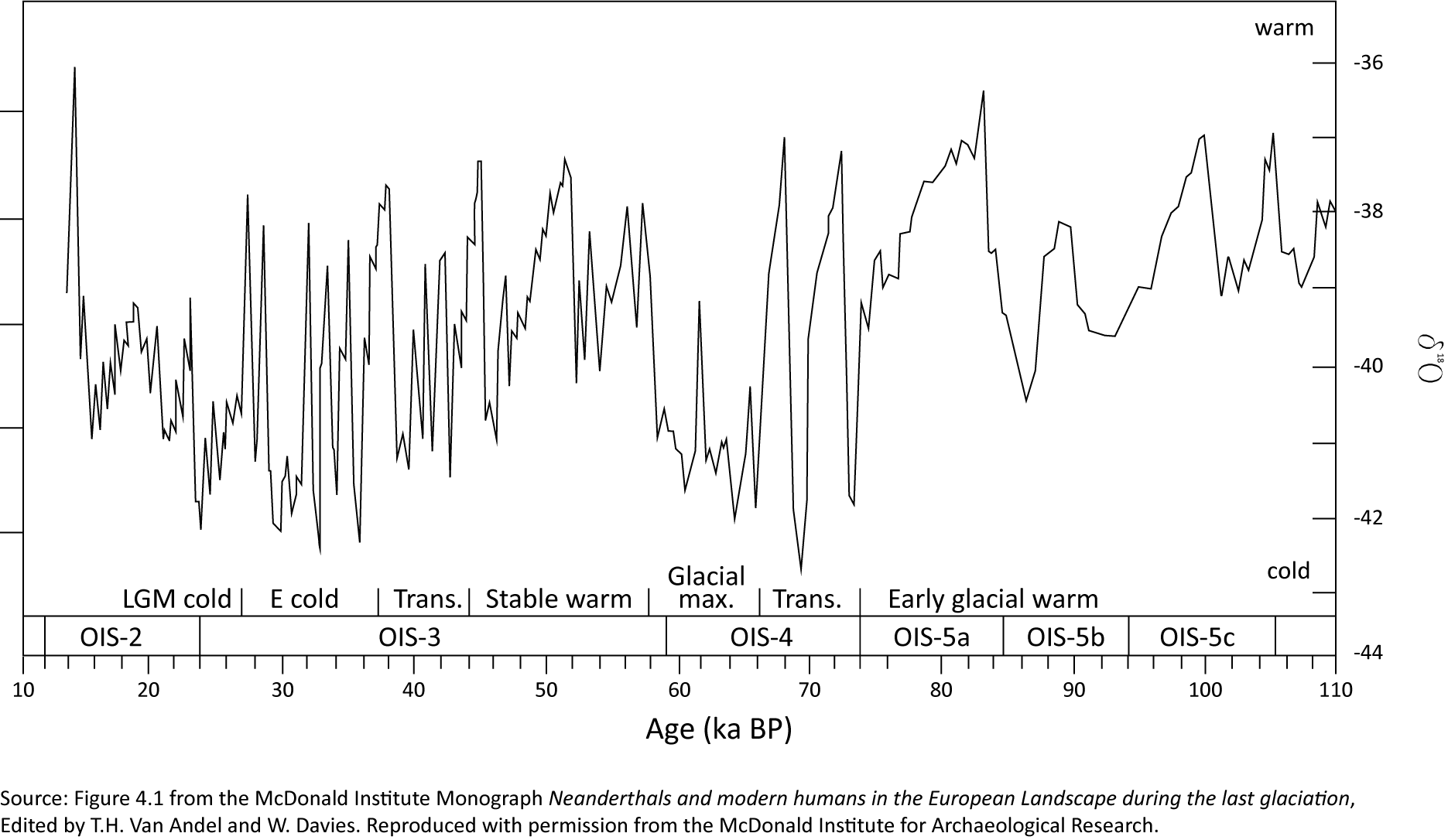
Undifferentiated Lower Ouse T1–T2 terrace deposits of Middle–Late Devensian Age have been recorded at Earith, Cambridgeshire (Bell 1970), where organic sediments in sandy gravels of the Hemingford Member have been radiocarbon dated to the Middle Devensian at c. 42140 +1890/–1530 (Birm88). At Woolpack Farm, Cambridgeshire, sandy gravels for undifferentiated T1–T2 terrace deposits were assigned on the basis of molluscan biostratigraphy and vertebrate remains to the Hemingford and St Ives members of the Ouse Valley formation (Gao et al. 2000). Terrace deposits of Middle–Late Devensian age were also recorded for the nearby Cam Valley as members of the Cam Valley Formation, with radiocarbon dates for the Sidgwick Avenue (T2) and Barnwell Station (T1) members of 37746±420 (Wk9388) and 19500±650 (Q-590), respectively (Godwin and Willis 1964; Wymer 1985, 184–187; Boreham 2013). The Sidgewick Avenue T2 terrace deposits have been stratigraphically correlated with the T1–T2 deposits of the Hemingford Member of the Ouse Valley Formation, and the T1 terrace deposits at Barnwell Station with those of the Ouse Valley's St Ives Member (Boreham et al. 2010; Gao and Boreham 2011, fig. 3).
The pollen, plant macrofossil and insect assemblages from Association B deposits yielded evidence of a treeless grassland with a temperate continental climate characterised by warm summers (16–17℃) and cool–cold winters (–11° to 4℃). These environmental proxies indicated a predominately open floodplain landscape, with taxon diversity suggesting a number of different plant communities and a landscape occupied by a mosaic of four major habitat types related to floodplain topography, sediment make-up and hydrology. These habitats comprised pond and marsh–fen, grassland, tall–shrub willow scrub, and disturbed ground plants and plant communities (Redman 1982; West 2000; Miall 2006).
Pond and marsh–fen habitats would have occupied the lower parts of the floodplain, with shallow water communities (Hippuris vulgaris, Zannichellia palustris) and damp–wet sedge marsh–fens (Carex spp., Sparganium erectum, cf. Filipendula, etc.). Grassland habitats with rich meadow and pasture grass–herb communities (Helianthemum, Sanguisorba officinalis, Poaceae, etc.) were likely to have occurred on floodplain flats and river terraces. Tall–shrub willow scrub communities (Salix, Poaceae, Rumex cf. acetosa, etc.) would have also occupied floodplain flats and river terraces. Disturbed ground plants and plant communities (Cirsium type, Plantago cf. major/media, Artemisia type, etc.) would have been present in floodplain flats and river terrace habitats and largely related to local erosion and sediment deposition conditions. Dung and phytophagous Coleoptera (Acrossus rufipes, Platystethus arenarius, Graptus triguttatus, etc.) also suggested pasture and dung communities for grassland and related habitats, and the presence of significant populations of large herbivores. Vertebrate remains for these herbivores included mammoth (Mammuthus primigenius) woolly rhinoceros (Coelodonta antiquitatis), bison (Bison priscus), horse (Equus ferus) and reindeer (Rangifer tarandus) (Stuart 1982; Bliss and Richards 1982). The eggs of intestinal parasites provided further evidence for the presence of large herbivores.
Local habitat types suggested by section samples comprised shallow floodplain ponds (⋜1.0m deep) with rooted aquatic plants and insects (Zannichellia palustris, Hippuris vulgaris, Helophorus grandis, etc.), surrounded by sedge marsh–fens with grassy meadows and pasture beyond (Carex spp., Sparganium erectum) (sections SS02, SS05), and marginal wet–damp grassy meadow (Carex spp., Sanguisorba officinalis, Potentilla type) (section WkS06). Sedimentary features and microfabrics in thin-sections also provided evidence for periodic inundation by overbank flooding. Similar Middle Devensian pond, marsh–fen and grassland habitats have been suggested by pollen and plant macrofossil assemblages for Earith, Cambridgeshire, and Sidgwick Avenue, Cambridge (Lambert et al. 1963; Worssam and Taylor 1969, 115; Bell 1970; Bell and Dickson 1971). The occurrence of scarabaeid dung beetles (Aphodiinae spp.) at Earith also indicate a similar presence of large herbivores (Bell 1970, 372).
The Palaeolithic record for the extraction area comprised abraded and reworked lithic artefacts within younger river terrace deposits with no in situ primary context archaeological materials recorded for these deposits. Artefacts typically appeared to be fluvially abraded, with the occurrence of Lower and late Middle Palaeolithic tool types indicating a degree of reworking from older deposits and in line with the evidence for temperate arboreal pollen and the mixing of older and younger sediments suggested by OSL age inversions. The small size of the lithic assemblage and its multiperiod make-up indicated that it was likely a palimpsest-type assemblage and represented a group of reworked artefacts produced by fluvial erosion and sediment deposition rather than directly from human activities. OSL dates also suggested that the artefacts were not contemporary with the terrace deposits in which they occurred and represented a time-averaged collection of material documenting a human presence in the region during the Lower and late Middle Palaeolithic.
Particular thanks are due to Paul Sprague, Tom Scott and Clare Fleming of the A14 Integrated Delivery Team for their support despite the work pressures on them. A debt of gratitude is owed to Kevin Davies and Dia Davies of Walters and plant operators John 'Jock' Barclay and Darren Hickman for finding bone and tusks. Many thanks are owed to Tom Hodgson and Luke Jarvis of Headland Archaeology for their very hard work throughout their time on-site. Special thanks are also due to Tony Walsh formerly Headland Archaeology and Highways Archaeologist Steve Sherlock for their support and help throughout the fieldwork and to Kasia Gdanic of the Cambridgeshire Historic Environment Team for her interest in the work. Rachel Wood of ORAU is thanked for her advice regarding the dating of sediment fractions.
A special debt of gratitude is also owed to Rob Batchelor and the rest of the post-excavation team for their very hard work in meeting various deadlines and for sorting out environmental stuff. Carla Ardis and James Ladocha are mentioned in dispatches, Carla for her work on the illustrations and James for putting the section photographs together. Review of the draft text was done by Chris Green.
Enid Allison would like to thank Phil Buckland who very kindly provided the MCR data using the BugsCEP program. Scott Elias and Harry Kenward are also thanked for their assistance with the identification of some 'problem' specimens.
Many thanks are owed to Matthew Davies and Emma Jarman of the McDonald Institute for Archaeological Research, Cambridge and to William Davies and Kate Pretty for their permission to use Figure 4.1 from the McDonald Institute Monograph Neanderthals and Modern Humans in the European Landscape during the Last Glaciation edited by T.H. Van Andel and W. Davies (1999).
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Terms and Conditions | Legal Statements | Privacy Policy | Cookies Policy | Citing Internet Archaeology
Internet Archaeology content is preserved for the long term with the Archaeology Data Service (ROR). Help sustain and support open access publication by donating to our Open Access Archaeology Fund.
 Home
Home