Cite this as: Johnstone, C. and Albarella, U. 2015, The Late Iron Age and Romano-British mammal and bird bone assemblage from Elms Farm, Heybridge, Essex, in M. Atkinson and S.J. Preston Heybridge: A Late Iron Age and Roman Settlement, Excavations at Elms Farm 1993-5, Internet Archaeology 40. http://dx.doi.org/10.11141/ia.40.1.albarella
A large animal bone assemblage was recovered from Late Iron Age and Romano-British deposits from Elms Farm, Heybridge, Essex. The site comprised an Iron Age settlement (Period 2) and its continuation as a Romano-British small town (Periods 3-5), with occupation extending into the Early Saxon period (6). Areas of the site were related to the public open spaces, domestic activities, small-scale industrial processes and a religious complex. A total of 9064 fragments were recorded (countable and non-countable elements) 8376 of which were recovered by hand-collection and the rest by bulk sieving. The bones were mostly well preserved, but a large proportion showed evidence of fresh breakage. Species present included most domestic animals together with a few wild mammals and birds. In all periods cattle bones dominated the assemblage, forming up to 90% of the fragments (80% using MNI).
The skeletal element distributions showed that all three main domestic species were being consumed and probably butchered on site with no specialised waste disposal patterns. Differences in the areas of the site used for rubbish disposal through time were detected. Butchery of the cattle bones was extensive and followed a typically Roman pattern. Mortality profiles showed that cattle were mainly used for traction, sheep for meat and secondary products such as wool, and pig for meat. Some changes in the age-at-death data were detected between Periods 3 and 4 (Early and mid-Roman period). Pathologies noted on the cattle metapodials and phalanges confirm the possibility that the animals may have been primarily traction beasts. The biometry of all domestic species indicates that improvements in the size of livestock took place in the Roman period. For cattle this took place immediately after the conquest, while the other species were improved around a century later (Period 4).
The process of Romanisation at Heybridge seems to have taken place in two stages; the increase in cattle size with the physical changes to the settlement immediately post-conquest and the changes in size of the other animals and changes to the underlying economy taking place a century later. Deposits associated with the temple precinct at Heybridge have yielded information on sacrifices made at the temple complex. The results from Heybridge fit well into the picture from other sites in the region and also from further afield, but also provide important new information on the use of animals during the Late Iron Age to Early Saxon times.
The site at Elms Farm, Heybridge, Essex, was excavated by Essex County Council Field Archaeology Unit (under the direction of Mark Atkinson) during 1993 and 1994-5. For a full description of the site and the areas excavated see Section 1 and for the site chronology see the Site Narrative.
The material considered in this report came from deposits dated to Periods 2 to 6 as very little bone was recovered from Periods 1 and 7.
Atkinson and Preston (1998) give a broad chronological overview in their interim report. The text that follows is a summary of that discussion. Features dated to Period 2 were very difficult to tie down to pre- or post-conquest, therefore the material they produced had in most cases to be combined in a single period. However, a number of definite Iron Age features were identified including the earliest structures associated with religious activities in Area J, possibly trackways underlying the Roman roads and many pits in northern areas of the site. The transitional period was one of widespread change at Heybridge, most obviously seen in the creation of an ordered settlement with metalled roads, regularly laid out plots of land and a focal area around the 'upgraded' religious complex.
Period 3 saw the continuation of the transitional period and the development of a number of distinct aspects of the settlement. These included the zonation of established activities, the upkeep and repair of roads and increased use of the pit area along the south of the site. They also included further development of the temple precinct and complex, the emergence of a public space (possibly market area or connected to the neighbouring temple complex) and the continuation of domestic activity across the northern half of the site.
The economic growth and prosperity of the settlement begins to take a downturn at the end of Period 3 and is more obvious during Period 4. Wide-scale dumping of silt was used in an attempt to raise ground level, possibly following a period of flooding. This caused a change in the use of areas, particularly Area I where the roundhouses were demolished and replaced by an open area, and by the realignment of ditches all over the site. The temple complex was also altered again, and although the circular structure became bigger the others were demolished.
Changes continued through Period 5 including a continuation of the marked decrease in activity on the outer edges of the settlement. The religious complex continued to be a focus of activity with new internal structures being built and changed, however the temple precinct was being encroached upon by domestic activity particularly pit digging along the boundaries. By the latest Roman period (Period 6) the temple precinct may well have been abandoned as the amount of domestic and even industrial activity encroaching on the area increased dramatically. The road system also seems to have been abandoned as buildings, pits and post-hole lines encroached and even blocked the roads. The Early Saxon period is quite poorly represented, and seems to be restricted to the outer areas of the settlement.
Within the framework of the updated Project Design (Atkinson and Preston 1997) there are several general and specific objectives to which the zooarchaeological evidence can contribute. The overall aim is to develop an understanding of the morphology and function of the settlement from Late Iron Age to Late Roman periods. Within this, the zooarchaeological analysis can contribute to the following specific objectives:
Almost all the bone material (240 of 249 boxes) recovered from Elms Farm was excavated by hand and is termed 'hand collected (HC)' in this report. Material collected during machine stripping was generally poorly stratified and hence not included in this study. In addition to the hand-collected material, a further quantity of bone was recovered from the residues of 618 sieved samples. These were termed 'bulk-sieved (BS) samples'. These samples were not necessarily taken as 'whole earth' samples; in other words, bones were occasionally hand-collected prior to the processing of samples. The volume of each sample varied and depended on the size of specific features; however, around 20 litres was the average. These samples were then floated and bone recovered from their residues. Only this sampling programme was undertaken, with no large-scale coarse sieving employed on site.
Vertebrate material was recorded directly into a computer using Paradox software. The database consists of a purpose-built series of inter-linked forms and tables.
Records of preservation, angularity (appearance of broken edges) and fragmentation were made for each context using various categories. Preservation records were only made for contexts containing 'countable' fragments (see below). For the context to be recorded as one category more than 80% of the fragments had to be of that category, mixed contexts were recorded as 'variable'. Categories for preservation were 'excellent', 'good', 'fair', and 'poor' and those for angularity were 'spiky', 'battered' and 'rounded' (Dobney et al. 1999). Fragmentation was recorded using the percentage of fragments in the following size categories: less than 4cm, 4-10, 10-20 and greater than 20cm. The percentage categories were: none, 0-10%, 10-20, 20-50 and greater than 50%. Fresh breakage was also recorded using these percentage categories. In addition, notes were made as necessary on the preservation, angularity and any other interesting feature of the context as a whole. For small contexts (less than fifteen fragments total) only the preservation was recorded.
Identification of most of the material was carried out using the reference collection of the Birmingham Zooarchaeology Laboratory (BZL, University of Birmingham). Some closely related taxa are difficult to distinguish and for this study only the following elements were used for separation, and are those for which the criteria seem most reliable. For caprines the lower deciduous 3rd and 4th premolars (Payne 1985), distal humerus, distal metapodials, astragalus and calcaneum (Boessneck 1969) and distal tibia (Kratochvil 1969) were used. Horncores were also identified but not used for quantification. For equids the shape of the enamel folds was used to differentiate species (Davis 1980). However, no complete or subcomplete molar rows were present in this assemblage to allow this to be undertaken; hence bones and teeth recorded as 'Equid' could contain species other than the domestic horse. In an attempt to separate the species the biometrical method of Davis (pers. comm.) was undertaken on the 1st phalanx. Differentiation of the domestic fowl, pheasant (Phasianus colchicus) and guinea fowl (Numida meleagris) was undertaken on all recorded elements (see below). Swan species were separated by morphological differences on the sternum (recorded as a non-countable element). Only the pelves of amphibians were used to distinguish frog from toad.
For the hand-collected material a selective recording system was used, as outlined in Davis (1992) and Albarella and Davis (1994), with minor modifications to suit this assemblage and to allow direct computer input. In brief, the following parts of the mammalian skeleton were always recorded and used for quantification: mandibles (with at least one tooth), loose mandibular teeth (at least half of the occlusal surface), cranium (zygomatic), atlas, axis, scapula (glenoid area), distal humerus, distal radius, proximal ulna, carpal 3 (2+3 in some species), distal metacarpal, pelvis (ischial part of acetabulum), distal femur, distal tibia, astragalus, calcaneum, distal metatarsal, proximal phalanges 1, 2 and 3. In order to avoid multiple counting, at least 50% of a given part had to be present. Single condyles of cattle, caprine and cervid metapodials were counted as halves, as were the central pig metapodials (lateral metapodials and phalanges were not recorded). Similarly the following parts of the avian skeleton were recorded: scapula (articular end), proximal coracoid, distal humerus, proximal ulna, proximal carpometacarpus, distal femur, distal tibiotarsus, distal tarsometatarsus. The following amphibian and small mammal elements were also recorded; amphibian scapula, humerus, radioulna, pelvis (acetabulum), femur and tibiofibula; small mammal, cranium, maxilla and mandible (M1 present), loose first molar, humerus, ulna, femur and tibia.
Horncores and antlers (with a complete transverse section) together with any non-countable elements from rarer species or parts displaying butchery marks or pathological conditions of interest were recorded but not used for quantification. In addition, numbers of cervical, thoracic and lumbar vertebrae were recorded for each context, by species (or species group) where possible or recorded as large mammal (cattle, horse, large cervid), medium-sized mammal 1 (caprine, pig, small cervid), medium-sized mammal 2 (dog, cat, hare), small mammal, or bird. Ribs with the articular ends present were also recorded for large- and medium-sized mammals. The weight of the bones was recorded by species for each context. Weights were collected as a means of assessing the fragmentation of the material (in conjunction with fragment numbers), rather than as a means of quantification.
Material from sieved samples was recorded following the same criteria described above. The number of identified specimens (NISP) was calculated for all taxa and the minimum numbers of individuals (MNI) were only calculated for the most common taxa. The MNI was calculated by simply dividing the total number of fragments of each element by the number present in the body. This was facilitated by the recording system containing only non-repeatable fragments.
Information on the presence of gnawing and burning, and details of butchery and pathology were recorded for each bone. Butchery marks were described by type (i.e. chop, knife mark, hook damage, sawing) and their position noted (using the diagnostic zones of Dobney and Rielly 1988), together with the orientation (using standard anatomical orientation). Pathologies were categorised, the position noted (as for butchery) and a detailed description made where necessary.
Sexing using morphological characteristics was only undertaken for pigs and chickens. The shape of the pig canines (and their alveoli) was used to differentiate boars from sows, whilst the presence of a spur on the chicken tarsometatarsus was used to distinguish cocks (and capons) from hens. With the chickens, this criterion is not totally reliable as females occasionally develop a spur and young-adult chickens may have bones that look osteologically mature (the ends are not porous) but have not yet developed a spur.
Information on the state of epiphyseal fusion was collected for all long bones and was recorded using a number of categories: fused, fusing (fusion line still visible), unfused (epiphysis, metaphysis or both) or juvenile (unfused and porous). If the bone was recorded as juvenile then a note would be made in the comments box if it was also considered to be neonatal i.e. very porous and very small. Bird bones were recorded as adult or juvenile. Analysis of fusion data was undertaken using the categories of O'Connor (1989).
Mandibular tooth wear stages were recorded from the dP4, P4, M1, M2 and M3, for both loose teeth and those in mandibles. Cattle and pig teeth were recorded after Grant (1982), while for caprine teeth the criteria of Payne (1973; 1987) were followed. Cattle and pig mandibles were assigned to the general age categories outlined by O'Connor (1989) and caprine mandibles to those of Payne (1973).
A complete list of the individual measurements taken on material from Elms Farm is given in Appendix 1 in the archive. Measurements mostly follow von den Driesch (1976), with the exceptions noted below. Measurements of cattle and caprine teeth were the maximum width (or length) of the given tooth (extraction of the teeth from mandibles was sometimes necessary to obtain the maximum). Measurements of equid cheekteeth follow Davis (1987). Pig tooth measurements follow Payne and Bull (1988) with the addition of the width of the central (i.e. second) cusp of the 3rd molar.
Humerus HTC and BT and Tibia Bd, for all species, were taken in the way described by Payne and Bull (1988) for pigs. Tibia SD for cattle, caprine and pig bones was taken in the anterior-posterior plane not the medio-lateral plane shown in von den Driesch. Caprine calcaneum measurements C and C+D were taken as indicated in Dobney et al. (c1996). Measurements on cattle, caprine and cervid metapodials follow Davis (1992). WMax and WMin were the largest and smallest diameters at the base of horncores and antlers, BC was the basal circumference and GL was the greatest dorsal distance (in a straight line) from the base to tip of a horncore.
Withers heights for horses were estimated using the criteria of Kiesewalter (as quoted in von den Driesch and Boessneck 1974) and those of Vitt (1952). For dogs, the withers heights and cranial indices were calculated following the criteria of Harcourt (1974). The log ratio technique of Simpson et al. (1960) was used to look at size variation in cattle, pigs, caprines and horses where there were insufficient numbers of individual measurements. The standard used for the cattle was the mean of the measurements from Period 2 from this site and for pigs was the mean of the sample from late Neolithic Durrington Walls (Albarella and Payne 1993). The standard for caprines was the mean of a sample of unimproved Shetland ewes (Davis 1996) and for horses the mean of a sample of Roman horses in Britain (Johnstone 1996).
Statistical tests were employed to determine the significance of observed differences in data. These have been employed in a slightly unorthodox way because of the limitations of archaeological data. For instance, in some cases there is no guarantee that specimens from a sample are completely independent. For the biometrical data the Student's t-test was used to determine the difference between two sets of data. This was undertaken in Microsoft Excel using the 't-test assuming equal variance' function. We assumed equal variance for all the data, as in our experience, the variance of two sets of data of the same measurement from the same element is unlikely to be significantly different. The test was only undertaken when the sample size was greater than ten.
A total of 249 boxes (34x46x15cm) contained the animal bone recovered from excavations at Elms Farm. Of these, 240 boxes contained hand-collected animal bone and the remaining nine contained the vertebrate material recovered by sieving. Animal bone was recovered from a total of 2502 contexts, 283 of which were unphased and hence the bone from them was not recorded. In addition, 943 phased contexts contained no 'recordable' skeletal elements and so do not appear in the archive. This left a total of 1276 phased contexts with 'recordable' fragments, which were recorded as described above. Of the total of 618 sieved samples only the residues of 194 contained 'recordable' material.
A total of 9064 fragments were recorded, including the 'non-countable' elements. Most of the fragments (8376) were recovered by hand-collection. Table 117 shows which species were present in the hand-collected and/or sieved assemblages by period. Table 118 gives the numbers of fragments by species and by period for the hand-collected material ('countable' elements only) and Table 119 gives similar information for the sieved assemblage. A number of complete and part skeletons were recovered from a variety of contexts and details are given in Tables 120 and 121.
| Taxa | Period 2 | Period 3 | Period 4 | Period 4-5 | Period 5 | Period 5-6 | Period 6 | |
|---|---|---|---|---|---|---|---|---|
| Bos taurus | Cattle | 227(35:2) | ||||||
| Sus scrofa L. | Pig | 101(4:1) | 50(8:1) | 29(23:2) | ||||
| Equus sp. | Equid species | 39(33:2) | ||||||
| Canis familiaris | Dog | 8(3:1) | 41(20:3) | 52(34:4) | 73(67:6) | 15(4:1) | ||
Animal bone was recovered from all areas and periods of the site although the quantity of material varied considerably between periods and areas. Only material from stratified deposits that could be reliably phased was considered for this report. For the analysis stage of the project the number of different periods had to be reduced to a manageable level; this meant that inevitably some periods had to be combined. This was mostly restricted to the later periods where contexts phased as 4-5 or 5 have been considered together as Period 4-5, and similarly Periods 5-6 and 6 have been amalgamated as Period 5-6. This seemed the most sensible approach given the smaller quantity of material from these periods.
The question of residuality arises for all multi-period sites. In this report residuality is defined as 'the occurrence of material datable to an earlier period in a context belonging stratigraphically to a later period'. This problem is particularly difficult to tackle in zooarchaeology, as animal bones are not intrinsically datable. We must therefore rely on the information provided by datable finds, such as pottery and coins, associated with the bones. This is not an ideal practice as there are a number of reasons why pottery should not be regarded as an accurate proxy of residuality in bones, but at the moment we have no better system available. Attempts made in the past to estimate residuality on the basis of the preservation conditions of the bones (see Dobney et al. c.1996) have been unsuccessful, as they were in fact measuring re-deposition and not residuality.
In our assessment of the bone assemblage from Elms Farm (Albarella 1996) we discussed the residuality problem and recommended that all contexts that were regarded, on the basis of other finds and stratigraphic considerations, as at a high risk of residuality, should be excluded from the analysis. Inevitably, at assessment stage, when a proper analysis of the finds had not been carried out yet, the residuality risk of many contexts was unclear. It has, however, finally emerged that most of these contexts had low levels of residuality (Mark Atkinson, pers. comm.) and have therefore been included in this analysis, while contexts that already at assessment stage were regarded as at a high residuality risk have been left out. There is little doubt that some of the bones included in this study are in fact residual, but we are confident that they are not of a sufficient number to affect the main results of our analysis.
Issues relating to the preservation, fragmentation and taphonomy of bones from archaeological sites are many and varied and have been discussed at length elsewhere (Luff 1993; Lyman 1994). The main factors affecting the preservation and recovery of bones are pre-burial processes (butchery, disposal strategies, scavenger activity, weathering), post-burial processes (nature of burial environment, reworking of deposits) and excavation strategies (selective excavation, hand/machine recovery, sampling regimes). When any number of these processes are combined, their individual effects can be difficult to determine and the end result not easy to interpret. However, as these processes can be very important in the interpretation and understanding of a particular assemblage of animal bones, at least some attempt has to be made to identify the major factors involved on that particular site.
Studying the preservation of the recovered bone fragments can be useful in determining to what extent the burial conditions on the site have adversely affected the bones, across the whole site and also detecting variations within it. Preservation records can also be used to assess the degree to which individual deposits may have been subjected to reworking. Assessing the degree of fragmentation (in combination with other observations) of the bones can be useful in determining factors in all three of the main categories stated above. It can help determine if pre-depositional factors such as butchery and carnivore activity, redeposition of material or excavation techniques have played a part in the current appearance of the assemblage. Many varied methods have been employed in the past to assess both the preservation and fragmentation of a given assemblage and almost all are subjective to some degree. This causes great problems when comparing the work of different zooarchaeologists as two people's definitions of a well-preserved fragment can differ greatly. However, as nobody has yet come up with a definitive, workable solution to either the problem of subjectivity or inter-worker differences, the best that can be done is explain the methods and criteria employed as fully as possible (see methods section and explanations below).
Starting with the preservation, an attempt was made to locate any trends or differences in the preservation by period, area and context type. Locating any general trends between periods proved to be quite difficult as the variability of the contexts within each period was great and exceptions to the general appearance were always present. In spite of this a few overall statements on the preservation by periods can be made, Periods 2 and 3 show the worst overall preservation and Periods 4 to 5 the best. Although Period 2 has the worst preservation it is not badly preserved, with most contexts being described as fair and angularity as battered. The best-preserved periods have a majority of contexts with good preservation and spiky edges.
Analysis of the preservation records by area shows that four areas (F, P, Q and R) have very few contexts containing bone, but based on what was present, Area R had uniformly poor preservation and the others mostly fair and battered. Of those areas with more than twenty recorded contexts, Area D had the worst overall preservation, Areas N and K the best, and the rest in between, with H, E and J having slightly better preservation than the others. As with the analysis by period, there are exceptions to the general trend in all areas of the site.
Looking at the same data by context type a more varied picture appears. Ditch contexts appear to have the worst preservation overall and well fills the best. Ditch contexts had the highest proportion of poorly preserved material and rounded fragments although the majority were recorded as fair and battered. Well fills had uniformly good or excellent preservation, which is hardly surprising given the waterlogged nature of the most of these fills. What is more surprising is the fact that many fragments displayed battered edges, suggesting some degree of degradation prior to burial. Pit fills were the most variable context types with reasonably equal numbers of contexts recorded as good and spiky, fair and battered or variable. Other context types contained mostly battered fragments with fair or good preservation.
The preservation of a fragment can be affected by its size, density, pre-burial alterations, disposal practice, burial conditions and degree of reworking, which may combine in many different ways. Looking at the overall preservation of material from whole contexts should even out the differences caused by size and density, and given that across the site the burial conditions (in terms of soil chemistry) are likely to be roughly equal, others factors may play a more important role in distinguishing differences between periods, areas and context types. Butchery will be looked at in more detail later on, but is not thought to affect the preservation of fragments to any great degree when taken over a whole site. Carnivore gnawing is a discrete and characteristic alteration to the bones and as such can be assessed as a separate issue from the overall preservation of the assemblage. This suggests that the observed intra-site differences in preservation are most likely to be affected by weathering, disposal practice, specific burial environment and degree of reworking. It has been suggested elsewhere (Dobney et al. c1996) that the angularity of the broken surfaces of the fragments is a useful way of assessing the extent of reworking within a deposit. The degree of rounding of broken edges can be taken as a measure of the degree to which a particular fragment has been moved around once it has been discarded. However, it should be noted that it cannot distinguish between degradation prior to burial (trampling, weathering) and that caused by reworking of deposits.
Overall preservation (including the angularity) of the assemblage from Elms Farm was quite variable, although this variability was mostly between contexts rather than within them. A few contexts (9610, 9796, 9828 and 10586) were the exception to this picture and showed distinct groups of bones with different preservation states. It is suggested that it is these contexts that are most likely to contain reworked material. It seems that, overall, the material from Elms Farm has not been severely degraded either prior to burial or as a result of reworking, indicating that a relatively high proportion of individual contexts may contain material derived from primary deposits. Several articulated whole and part skeletons were recovered from a range of deposits (details in Table 120). Although most of these were not recognised as articulated remains at the time of excavation, their presence does suggest that the contexts in which they were found are primary deposits.
Several methods of assessing the degree of fragmentation have been used on the Elms Farm assemblage. The first looked at the general assessment of fragmentation as recorded in the preservation database (see methods section). This gave an overall picture of the level of fragmentation of the whole context. The second method involved calculating the average fragment weights by species for the domestic mammals. This method was used to assess the effect of fragmentation on different species. Other methods of studying fragmentation include looking at the ratio between isolated teeth and mandibles.
Looking at fragmentation by period, both methods show that only two periods stand out as substantially different, Periods 4-5 and 6, which show a higher proportion of larger fragments. In Period 4-5 this probably results from the presence of several articulated skeletons in one well deposit, but this is not the case in Period 6. The other periods are remarkably similar. However, the difference between the weight of cattle and horse fragments (assumed to be roughly equal weight originally) shows that cattle bone was quite heavily fragmented throughout all periods.
Fragmentation by context type shows a much more varied picture. The wells show higher proportion of larger fragments, as would be expected considering the articulated skeletons found in these contexts. The pattern is similar to that seen in Period 4-5 as the same factors are affecting the two categories. Post-holes, gullies and layers all show quite a high degree of fragmentation, with more than 50% of fragments in more than 70% of the contexts being between 4 and 10cm in size. This may reflect: 1) the size of the feature in the case of the post-holes - i.e. large fragments will not fit, and 2) the type of feature in the case of floors and layers - i.e. the material may have been heavily trampled and fragmented prior to burial. Surprisingly, the ditch contexts show a similar rather fragmented picture, which may suggest secondary deposition of material into these features, supported by the positive correlation between preservation and fragmentation in these contexts (see above). Pit fills show variability in the fragmentation but on the whole the fragments are larger than in other context types.
The average fragment weight by context type is also very variable and less easy to interpret, as some context types do not contain certain species. It does show once again that the degree of fragmentation of cattle bones is much higher than other species in relation to their size. It also confirms those cattle fragments in pits and wells were larger than in other contexts. More fluctuations are visible in the pig and sheep/goat fragments, with smaller fragments in floor contexts suggesting that even (initially) smaller bones were being heavily fragmented here.
The fragmentation by area is also very varied, but the central areas J, K and L tend to have larger fragments of all the main domesticates. Perhaps this reflects the fact that these areas are either or public or located in the vicinity of public spaces and hence that rubbish would be cleared quickly into pits or other appropriate features rather than being left around to be weathered or trampled.
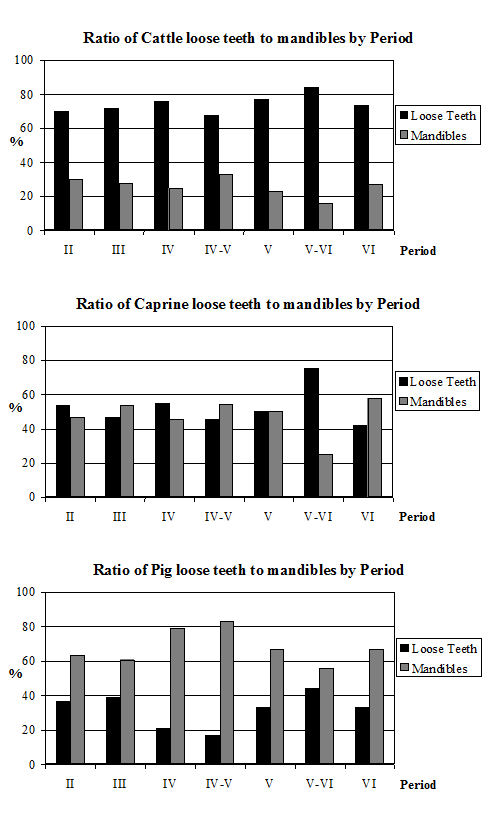
Analysis of the ratio of isolated teeth to mandibles of the cattle shows that in all periods there is a predominance of loose teeth (Figure 584), though this is slightly more pronounced in Period 5-6. Caprines show a nearly equal proportion of isolated teeth and mandibles in all periods except Period 5-6, where teeth dominate. Pig mandibles outnumber teeth in all periods and this is particularly pronounced in Periods 4 and 4-5. Although the evidence seems to indicate that cattle remains were much more affected by fragmentation than the other species, the higher proportion of cattle loose teeth can probably be explained by the fact that these teeth are larger and therefore more frequently recovered. Caprine and pig loose teeth are quite small and were probably frequently overlooked during the excavation. Although the quantity of loose teeth recovered by sieving is rather small (Figure 585), the marginally higher numbers of caprine and pig teeth tentatively confirms this suggestion.
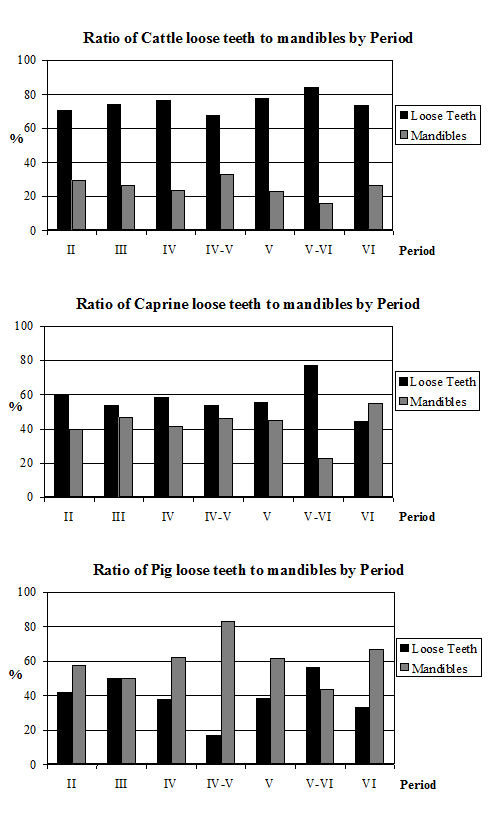
One of the major contributing factors in the fragmentation of this assemblage is the very high level of fresh breakage (the other being the high degree of butchery, which will be discussed in more detail below). In most periods more than 50% of the fragments in over 40% of the contexts were subject to fresh breakage (Figure 586). In many of these contexts the proportion of freshly broken fragments was nearer 80%. Significantly, not a single context was recorded as having no freshly broken fragments and material from only 10% of the contexts showed less than 10% fresh breakage.
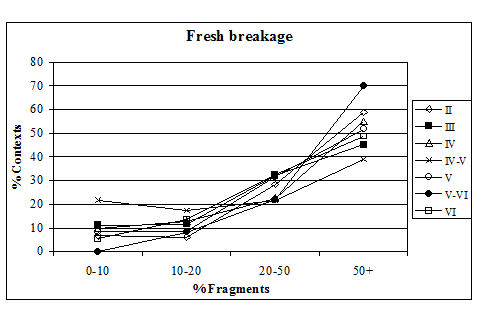
Fresh breakage was manifested as both clean breaks across fragments and also the degradation of edges and surfaces. All these types of fresh breakage suggest that a great deal of the vertebrate material was roughly treated during excavation, perhaps as a consequence of time pressure. This has unfortunately led to the loss of potentially useful data. Larger features were emptied using mattocks and shovels (Mark Atkinson, pers. comm.), which explains this phenomenon.
Evidence of scavenger gnawing was recorded by individual bone and records are only available for the 'countable' fraction of the assemblage. Context 6251 was noteworthy in that almost all the fragments were gnawed. As all the observed evidence of gnawing appeared to be characteristic of canids, it will henceforth be referred to as dog gnawing, this being the species most likely to be responsible for most, though not all, of the chewing. The overall level of dog gnawing was around 3.5% of the recorded fragments. Splitting this down by period, Periods 2 and 4-5 show higher levels of gnawed material (4.2% and 6.7% respectively). This level is relatively low and suggests that most of the bone material was buried quite rapidly after disposal or that dogs were unable to access the material.
Table 117 gives a complete list of species present in the Elms Farm assemblage together with information on the recovery method that produced those remains. The pattern that emerges from this table is that the bones of larger species are over-represented in the hand-collected material, whereas sieved samples are biased towards smaller taxa. This was expected, considering that the sieving was not carried out on 'whole earth' samples. Some of the species were only present as 'non-countable' fragments and are indicated as such in Tables 118 and 119, which give the numbers of individual skeletal parts (NISP) for the hand-collected and sieved material respectively. For the hand-collected material, as is the case on most archaeological sites (from the Iron Age onwards), the main domestic species, cattle, caprine and pig dominate the assemblage. Other domesticates such as horse, dog and chicken formed the next most numerous group of species and wild species such as deer and wild birds were the least numerous. The sieved material showed a similar pattern with the main domesticates making up most of the assemblage and the wild species forming the remainder. In terms of the range of species present, the Elms Farm assemblage is almost identical to that from Ivy Chimneys at Witham, also in Essex (Luff 1999).
A number of articulated whole and part skeletons were recovered from a range of deposits (details in Tables 120 and 121). Most of these were not recognised as articulated remains at the time of excavation, which has made the attribution of bones to specific individuals sometimes difficult.
Looking more specifically at the main domestic species, the minimum number of individuals (MNI) was calculated for cattle, sheep and pig in the main periods (Tables 122, 123 and 124). MNI calculations for all three domesticates show that teeth always provided the highest MNI, indicating that differential preservation of bones and teeth plays a major role in the skeletal element distribution patterns observed for Heybridge. The cattle MNI values range from 51 in Period 4-5 to 158 in Period 4, caprine values range from 13 in Period 4-5 to 87 in Period 3 and pig from 5 in Period 4-5 to 18 in Period 2.
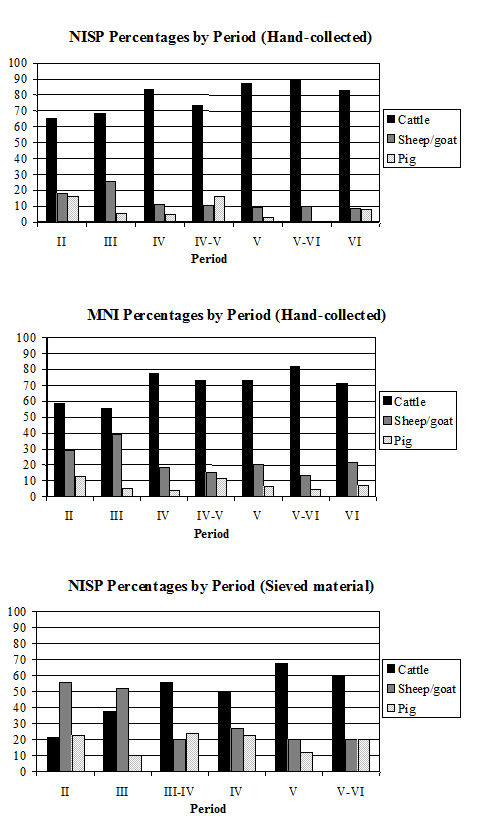
Comparing the relative frequencies of species by period for both the hand-collected and sieved material some clear differences can be observed. Firstly, comparing the NISP and MNI for the hand-collected material (Fig 587), it can be seen that the proportion of sheep/goat bones was higher using the MNI method. Therefore, the MNI method is more likely to reflect the true proportions of the species as it is less affected by taphonomic and recovery biases. This was particularly noticeable in Periods 2 and 3 for which the NISP of the caprines is higher than in other periods. Looking at the NISP for the sieved material (there were too few fragments for MNI calculations) it can be seen that in Periods 2 and 3 the proportion of sheep/goat is higher again, in fact overtaking the cattle. The proportion of cattle is reduced in the later periods but is still predominant. This indicates that there is a distinct bias towards the larger cattle fragments in the hand-collected assemblage. Therefore, if the sieving program had been more extensive and carried out using whole-earth samples, the overall proportions of species would probably have been different and would have reflected the true relationship more accurately.
While taking into account the recovery issues detailed above, there is still a clear predominance of cattle remains in the later periods (4 onwards) at Heybridge. Even in the early periods (2 and 3) the number of cattle remains is greater than the other species, although by not such a great margin. This pattern of high numbers of cattle in the Iron Age is a phenomenon typical of eastern England according to the study carried out by Hambleton (1999). In comparison with other sites in central England (including East Anglia and the Midlands up to the Welsh border) the Heybridge species proportions fit well within the overall trends in all periods (Table 125, Figures 588, 589, 590 and 591). Mostly the Heybridge figures are towards the edges of the distribution but are in no way outliers.
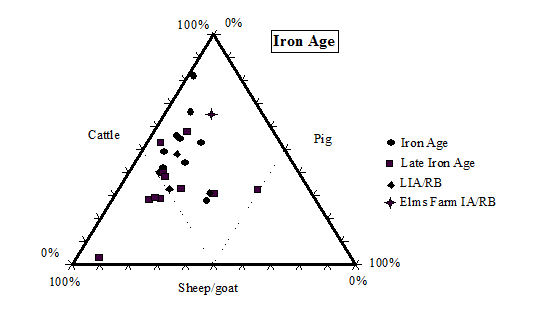
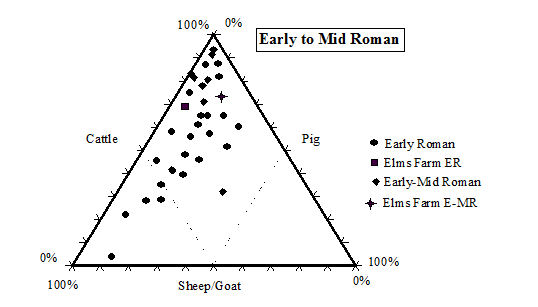
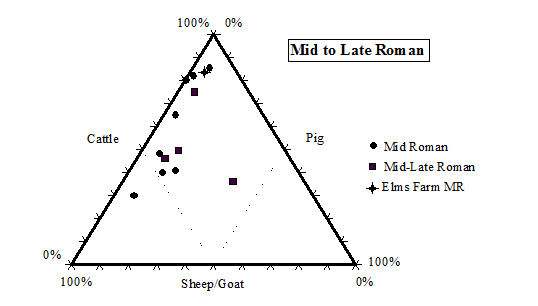
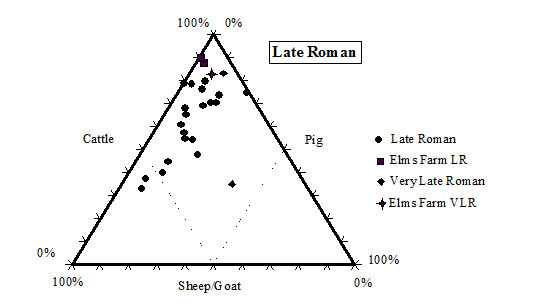
The main pattern that emerges is that cattle, taking into account its much larger bulk, would have provided by far the greatest amount of meat. The main reason for cattle breeding may not have been meat production (see ageing discussion below), but undoubtedly most animals would have eventually been used as food. The great importance of cattle for the Romano-British economy and diet is well known (see King 1978; 1984; 1999; and Grant 1989) and the data from the Elms Farm assemblage support this view. The typical Roman diet, as practised in Italy, would mainly have been based on pork consumption, but King (1978) suggests that the tradition of extensive beef consumption may have been imported to Britain by central European legions of the Roman army. This would explain the similarity in the distribution of species between military sites based in Britain and their equivalent in Germany and Gaul (see also King 1984).
King (1999, 180) has also suggested that military sites (likely to be more fully Romanised) would have greater proportions of cattle (and to a less extent pig) than rural civilian sites, which were more likely to continue the native Iron Age tradition. Urban settlements would be intermediate between the two. The sequence of site types with increasing proportions of cattle and pig suggested by King (1999, 180) is the following: 'rural settlements, villas, secondary urban centres, urban sites, military sites, legionary sites'. This is obviously a generalisation and we should not expect all sites to conform this trend. It is however, interesting to note that with its rather high (in comparison to other sites in central England) frequency of cattle, but relatively low proportion of pigs (Figure 589, Figure 590 and Figure 591), in the Roman periods the site of Heybridge has an intermediate status in King's sequence. Therefore, the site might well fit with the pattern described for 'secondary urban centres'. We have, however, to consider that the aforementioned long tradition of cattle breeding in eastern England and the likely poor rate of recovery at Elms Farm, also affect the relative frequency of the main domesticates.
With regard to changes over time, there is an expected and quite substantial increase in cattle frequency moving from the Late Iron Age to the post-conquest period, and a much more slight but steady increase going towards the end of the Roman period. The latest period, which may have a Saxon component, sees a slight increase in sheep numbers at the expense of cattle, which may indicate a decline of Roman influence and a partial return to a more traditional emphasis on sheep breeding. King (1999) has also observed this trend on other British sites.
Because of problems in determining, on archaeological grounds, whether some Period 2 deposits were pre- or post-conquest they were put into sub-periods in certain areas. Where the numbers of bones recovered was high enough for a meaningful analysis, slight differences between sub-periods 2A and 2B can be seen. The trend in Areas D, H and K show an increase in cattle numbers in 2B while the opposite is seen in Area L. This may suggest that in Areas D, H and K the Period 2A deposits are pre-conquest and 2B post-conquest. This is probably an oversimplification and cannot be taken as conclusive due to the relatively small numbers of bones involved.
The examination of the NISP of the species by context type (Table 126) reveals that pit fills contain more than half of the overall bone assemblage, followed by layers and ditch fills making up a further third of the assemblage. Wells provided the next highest quantity of bone followed by post-holes, floors and gullies. Other context types produced less than 100 fragments each. The proportions of the main domesticates in the context types (Figure 592) with more than 100 fragments shows the same overall pattern as the period analysis, with high numbers of cattle and 10-15% sheep/goat and smaller amounts of pig. This pattern alters in the floor deposits (contexts 16292 (G413), 6165 (G496), 6724 (G495), 7472 (G857) and 7572 (G857)) where nearly 100% of the fragments were of cattle. This may be a feature of the greater degree of fragmentation in these deposits, hence the relatively larger cattle fragments are the only ones that are recordable under protocol.
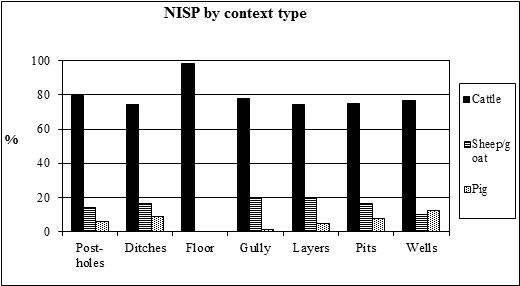
As the site was divided into areas probably at least loosely associated with function, it made sense to examine the species proportions within each area. The distribution of bones (Table 127) shows that the more central areas - H, I, J (the temple precinct), K and L - contributed the most, followed by the slightly more peripheral areas (D-G, M and N). The northern edges of the settlement and the hinterland areas produced the least bone. Examination of the areas from which over 500 fragments were recovered was undertaken by period. The NISP values indicate that rubbish disposal was concentrated in different areas during different periods (Figure 593), suggesting that the main occupation areas may also have shifted through time. In Period 2 most of the animal bones are concentrated in Areas H and L, and to a lesser extent in Areas D and K. During Period 3 bones are concentrated in Area I, the only period in which this area is extensively used for rubbish disposal. Although there are two distinct elements to this rubbish disposal, in pits and as spreads of material, there are no perceivable differences between the animal bones from the two context types. This suggests that the material in the pits may have been placed there after having been exposed for a period of time. Most bones from Period 4 are from Area H, with smaller concentrations in Areas J and K. The later periods show a concentration of discard activity in Area J, which agrees with other archaeological evidence, for example pit digging, suggesting that the temple precinct fell out of use at this time.
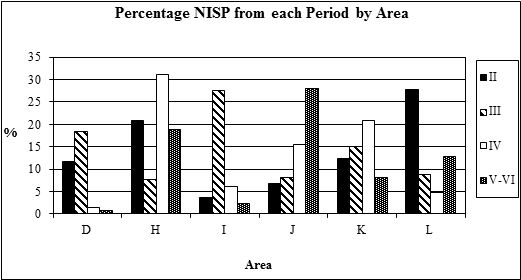
Analysis of the MNI data by area (Figure 594) shows that the general picture of high cattle numbers is seen across most periods of most areas. There are however, a few noteworthy exceptions. In Area D Period 4, Area K Period 2 and Area I Periods 3 and 4 the proportions of cattle and sheep are nearer to being equal. Area J shows a very high proportion of sheep/goat remains in Period 3, the significance of which is discussed in the sheep/goat section below.

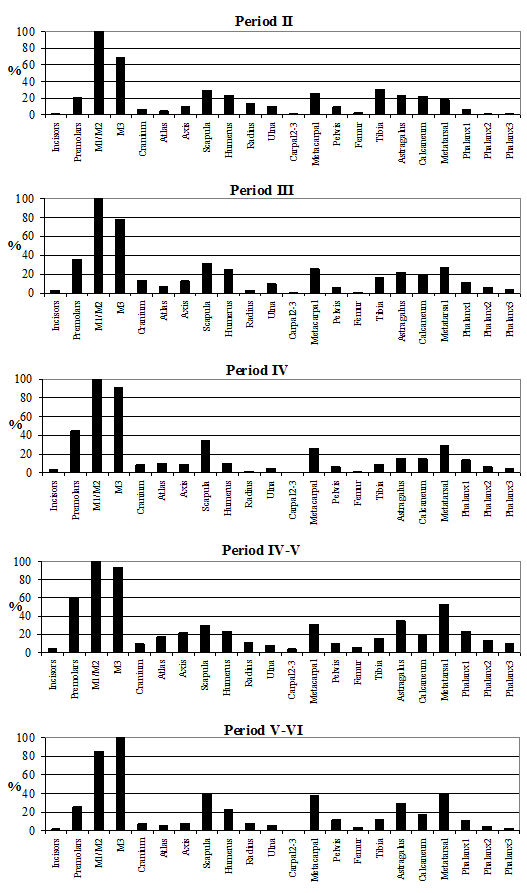
The skeletal element distribution is given in Table 122 and Figure 595. The frequencies of skeletal elements have been calculated using the MNI values rather than the NISP to eliminate the bias from elements that occur more frequently in the body. The graphs show the MNI; these were 85 in Period 2, 123 in Period 3, 158 in Period 4, 51 in Period 4-5 and 102 in Period 5-6. In all periods the highest MNI values were gained from permanent molars. The other teeth made up the next highest values in most periods. As discussed before this is a consequence of the differential preservation between bones and teeth. The distribution pattern of the post-cranial elements is similar between the periods. The most common elements were scapula, humerus, metacarpal, astragalus and metatarsal and the least common were generally the incisors, cranium, radius, carpal 2/3, femur and 2nd and 3rd phalanges. Horncores are not illustrated on the figures, as they are 'non-countable' elements; however, their NISP values are given in Table 122. Their relative percentage is around 20% for Period 3 onwards but is only around 5% for Period 2.
It appears that there were no specialised waste disposal patterns, as there are no peaks associated purely with primary butchery waste or food refuse. This suggests that animals may have been killed, butchered and eaten on the site and that no identifiable craft or industrial activities could be detected among the cattle bones. The pattern observed has more to do with preservation and recovery than with disposal practices. The smallest elements are consistently missing, suggesting that recovery bias is the factor here. Of the larger elements, the femur and radius are also consistently poorly represented, which is more likely to be a problem with preservation, as these are somewhat more fragile than other elements (remembering that for most elements only the distal end was recorded). The peaks correspond to the larger and more robust post-cranial elements and the teeth, corroborating the theory that recovery and preservation biases are responsible for the observed skeletal element distributions. The lack of horncores in Period 2 could either be a factor of the relatively poor preservation in this period or there could be a number of polled individuals present in the assemblage (although no direct evidence of this was noted).
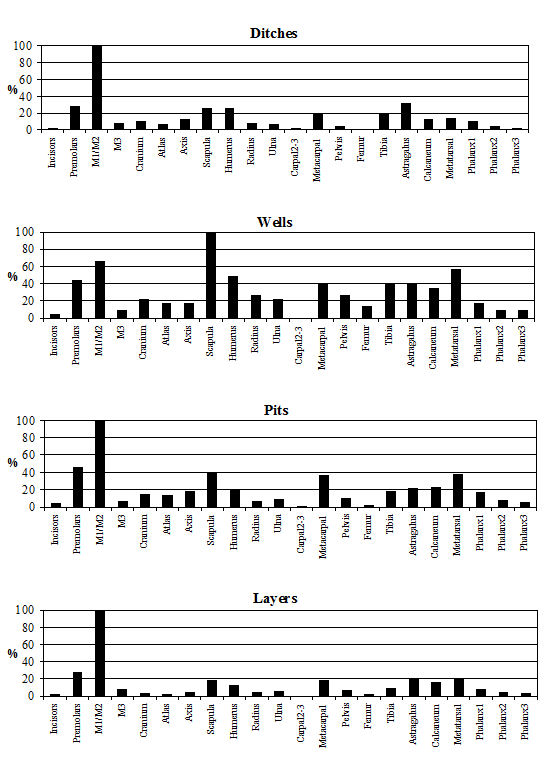
Analysis of the distribution of body parts by context type (Figure 596) shows that for ditches, pits and layers the observed pattern was similar to the distribution by period. Well deposits stand out for having a much greater proportion of scapulae and far fewer teeth. Analysis by area (Figure 597) shows that, while minor differences between areas are apparent, these differences are more likely to be the result of differential recovery and preservation rather than refuse disposal patterns. Areas K and N show a reduced proportion of teeth and a corresponding increase in post-cranial elements; this may reflect better preservation of post-cranial material or a real deficiency of teeth.
Analysis of the cattle vertebrae recorded in the non-countable section (Table 128) shows that the vertebrae are under-represented in all periods and that this is particularly true of the thoracic vertebrae.
| Cervical | Thoracic | Lumbar | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cattle | NISP | MNI | % | NISP | MNI | % | NISP | MNI | % | NISP | MNI |
| Period 2 | 7 | 2 | 50 | 6 | 1 | 25 | 4 | 1 | 25 | 17 | 4 |
| Period 3 | 17 | 4 | 36 | 24 | 2 | 18 | 25 | 5 | 46 | 66 | 11 |
| Period 4 | 28 | 6 | 32 | 49 | 4 | 21 | 51 | 9 | 47 | 128 | 19 |
| Period 4-5 | 24 | 5 | 42 | 41 | 4 | 33 | 13 | 3 | 25 | 78 | 12 |
| Period 5-6 | 20 | 4 | 40 | 18 | 2 | 20 | 21 | 4 | 40 | 59 | 10 |
| Total | 96 | 138 | 114 | 348 | |||||||
The cattle skeletal element distribution at Heybridge is similar to that observed at other sites in the area which have been interpreted as sites with no specialised activities, such as Braintree (Luff 1976 and Smoothy 1993), Caesaromagus (Luff 1992) and Colchester (Luff 1993). In contrast, sites with different skeletal distributions such as those at Harlow (Legge and Dorrington 1985) and Sheepen (Luff 1985) are interpreted respectively as a temple and an industrial site.
Butchery marks were prevalent on cattle bones, with an overall figure of 28% of bones displaying one or more marks. The lowest percentage of butchered bone was in Period 2 (14%), with the rest of the periods having around 30% of butchered cattle bones. The percentage for Period 2 at Heybridge is identical to the overall figure for Roman sites in the Netherlands (Lauwerier 1988), which is half that for the Roman levels at Heybridge. This may perhaps indicate that butchery practices at Heybridge were of a more intensive nature in most periods.
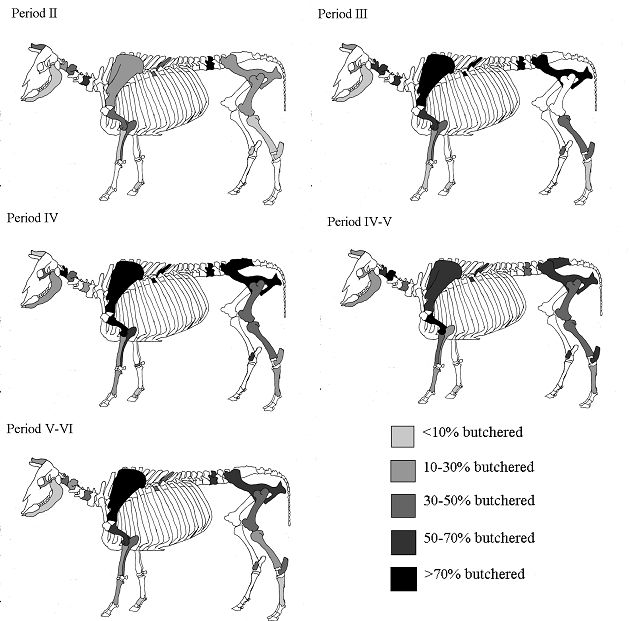
Figure 598 shows the percentage of bones displaying butchery marks. Calculating the percentage of bones with butchery marks helps to reduce the bias created by the selective recording scheme. For instance, under the selective recording scheme, the data are biased by indicating that all the butchery marks on the humeri were at the distal end whereas in fact that is the only part recorded. By calculating the percentage of bones with butchery marks that bias is eliminated. Calculating the percentage of bone butchered also allows direct comparison between periods where very different numbers of bones are present. Overall patterns of butchery on the cattle skeleton were consistent between the later periods but differ slightly from those seen in Period 2. These differences were mostly in the amount of butchered bones in relation to the quantity of bones recovered. The areas of the skeleton with the highest percentage of butchered bones were the cervical vertebrae, scapula glenoid, elbow joint, pelvic acetabulum and hock joint. These are the areas commonly used for the primary butchery of the whole animal into a carcass (removal of head and lower limbs) and its initial dismemberment into large joints.

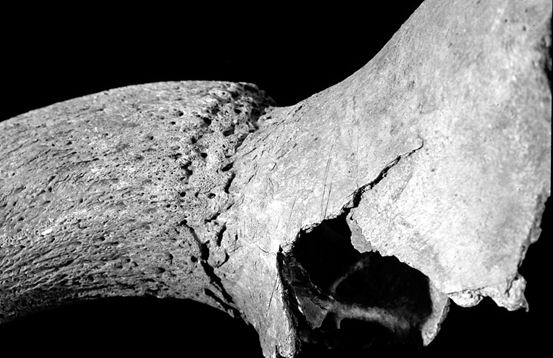
Most of the butchery marks were chops in the areas mentioned above, but several other types of butchery were noted including knife marks and hook damage. The term 'hook damage' is used to describe irregular holes in the blade of the scapula (Figure 599), which are thought to derive from hooks being put through the shoulder to hang it up for smoking or other curing processes (Schmid 1972). This damage was noted on many scapulae from Heybridge, particularly from the post-conquest periods. In conjunction with the hook damage, a particular pattern of chops was noted around the glenoid area of many of the scapulae (Figures 599 and 600). This consisted of one or more vertical chops removing the tuberosity, and slices off the rim of glenoid cavity medially, laterally and posteriorly. In addition the spine of some scapulae was also removed vertically. These specimens are more similar to what Dobney (2001, 41) regards as representing '“brined” and cold-smoked joints', than to the hooked scapulae with no trimming of the glenoid cavity, which were found in 4th-century AD levels at Lincoln and were thought to reflect hot-smoking with no immersion in brine (Dobney 2001, 41).
Other frequently repeated butchery patterns are also shown in Figure 600 for the humerus and astragalus. These kinds of butchery have been noted on many Roman assemblages (described in Maltby 1989), in particular the patterns illustrated at Lincoln (Dobney et al. c1996) and the Netherlands (Lauwerier 1988) are very similar to those found at Heybridge.
Although the overall pattern of butchery of the carcass shows no particularly unusual traits, there were patterns of butchery on specific elements that are highly specialised. The distribution and orientation of most of the chop marks form a distinctively Roman pattern of butchery that has been observed on many sites and intensively analysed by Maltby (1989). This pattern is characterised by intensive chopping of the bones, mostly beyond what would be needed for primary butchery and jointing of the carcass. These include marks consistent with the removal of the tongue, removal of the horncores (Figure 601), marrow extraction and production of broth (see below). Maltby has shown that this pattern is indicative of specialist, large-scale butchers operating in the vicinity, who were utilising the carcasses to their fullest extent. Maltby saw the Roman butchery pattern in urban and military assemblages but very infrequently in rural ones, suggesting that only settlements above a certain population size needed a full-time specialist butcher. The fact that the Roman butchery pattern is seen at Heybridge suggests that the population was large enough to need the services of such a specialist.

Several contexts (5472 (G436), 10090 (G830), 13407 (G626) and 14492 (G703) contained a large proportion of highly butchered fragments (Figure 602). The material from these contexts showed the butchery patterns described above, but other elements were also affected and the fragmentation was much greater. Most articular ends and smaller bones were chopped into small pieces and the shafts were also heavily fragmented. Similar deposits have been noted from several Roman sites and have been termed 'soup-kitchen' deposits (Van Mensch 1974). Van Mensch's paper has photographs of fragments displaying butchery patterns that appear almost identical to those found in the soup-kitchen deposits at Heybridge. Ethnographic parallels (Saint-Germain 1997) have been cited for the practice of boiling bones to extract fat and other dietary elements in prehistoric communities where the need to obtain the maximum nutritional value from one animal was paramount. However, this is unlikely to have been the case in Roman times. It may be that this represents extraction of fat from bones for semi-industrial or craft purposes, as the fat obtained in this manner is much purer than that recovered from marrow. The word 'soup' and its consumption is thought to be a Germanic custom (Van Mensch 1974), so possibly areas in which these deposits are found may indicate the presence of people of Germanic origin, perhaps ex-soldiers and their families. Either way, these bones indicate a use beyond the processing of meat for ordinary consumption.
We are at present collaborating with researchers in an attempt to detect whether the bones from soup-kitchen deposits had been cooked (Roberts et al. 2002).
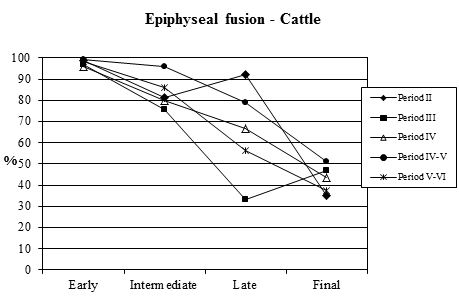
Examining the data concerning epiphyseal fusion first (Table 129, Figure 603), the proportion of cattle in all periods that had reached skeletal maturity before death was high. Over half the bones in the late fusing category were fused (except Period 3) and over a third of all vertebrae (final category) were also fused. This suggests that, on the fusion evidence, over a third of the cattle from Heybridge were mature individuals, probably more than five years old (data from Silver 1969), and that very few (less than 4%) were less than one year old. The pattern seen at Heybridge is also seen at several other Roman sites, such as Lincoln and Exeter. This suggests that in all periods there was no intensive exploitation of these animals for beef or for dairying but possibly the cattle were used as traction beasts with beef production a secondary consideration. However, we need to look at the dental evidence in order to draw accurate conclusions on the cattle kill-off pattern.
| Cattle | Early | Intermediate | Late | Final | |
|---|---|---|---|---|---|
| Period 2 | Fused | 78 | 109 | 24 | 6 |
| Unfused | 1 | 25 | 2 | 11 | |
| % fused | 99 | 81 | 92 | 35 | |
| Period 3 | Fused | 199 | 112 | 2 | 31 |
| Unfused | 5 | 36 | 4 | 35 | |
| % fused | 97 | 76 | 33 | 47 | |
| Period 4 | Fused | 267 | 167 | 4 | 56 |
| Unfused | 11 | 42 | 2 | 72 | |
| % fused | 96 | 80 | 67 | 44 | |
| Period 4-5 | Fused | 164 | 93 | 15 | 44 |
| Unfused | 1 | 4 | 4 | 42 | |
| % fused | 99 | 96 | 79 | 51 | |
| Period 5-6 | Fused | 157 | 148 | 13 | 22 |
| Unfused | 3 | 24 | 10 | 37 | |
| % fused | 98 | 86 | 57 | 37 | |
Tooth wear stage data were recorded using the wear stages of Grant (1982) but her method of calculating the mandible wear has a number of problems associated with it which have been discussed elsewhere (O'Connor 1988). In view of these difficulties, the broad age categories of O'Connor have been employed for the analysis of the cattle data. Tables 130 and 131 give the data from cattle mandibles and individual teeth. These confirm the epiphyseal fusion data and show that very few of the cattle remains at Heybridge were from young individuals. In fact no mandibles from neonatal or juvenile individuals and only twenty-two immature mandibles, representing 4% of the total number, were recovered. Almost all the mandibles were aged to the subadult and adult categories with at least a few elderly individuals in all periods. Preservation will bias against very young and immature individuals as the bone is more fragile and the teeth are more likely to fall out of the mandibles. For this reason the loose teeth were examined (Table 131) to see if there was evidence of bias against younger individuals in the mandible data. As this did not appear to be the case, we can assume there is a real dearth of younger animals.
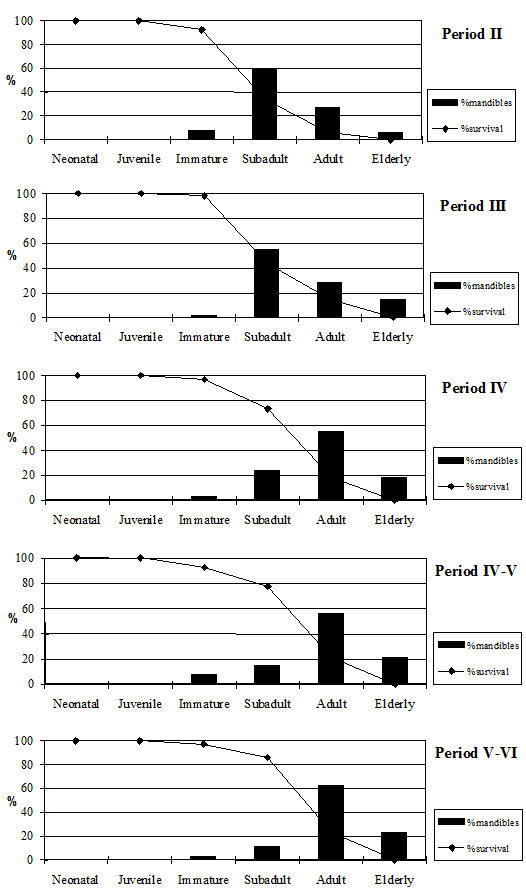
There are, however, differences between periods. Periods 2 and 3 have very similar patterns (Figure 604) with most individuals reaching the subadult category and then being killed off, with substantially fewer individuals reaching the adult and elderly categories. This pattern is what would be expected from a meat-based economy with most animals being killed when their maximum body weight has been reached and a few older individuals kept for breeding and traction. The pattern changes in Period 4, with most animals living until the adult category and an increased number of elderly animals. The later periods continue to show this pattern. This suggests that the cattle economy in these periods was less driven by meat production, and more animals were being kept for traction. It is also possible that in the later period a greater number of bullocks were sold to the market, perhaps to feed larger urban centres or military settlements. This hypothetical move towards an increasing production role of the site is, however, not supported by any other evidence, and we must therefore regard the change in husbandry strategy as a more likely explanation.
Other British sites, including Exeter (Maltby 1979) and Colchester (Luff 1993), indicate that in the Iron Age cattle were used as a multi-purpose beast, whereas the Roman economy was heavily based on beef production. However, on both these sites the deposits yielding this information could have been influenced by the presence of the army. Sites with a more civilian nature, such as the coloniae of York (O'Connor 1988) and Lincoln (Dobney et al. c1996) and several sites in the Netherlands (Lauwerier 1988), show a similar trend to that found at Heybridge. Many of the sites listed in Table 125 also show a predominance of adult cattle individuals. This suggests that although the military probably required some young beef, the majority of civilians in Britain reared cattle mainly as working animals. This strategy not only occurred throughout the Roman period but even seems to have been more greatly intensified with the increasing Romanisation of the region.
Only a few cases of oral pathologies were noted. These included a single case of hypoplasia (an incisor, context 4925, Group 739, Period 4) a condition generally related to nutritional or other environmental stress. Two teeth (contexts 6283, Group 541, and 10287, Group 303) were noted with either congenital or growth defects. Ten mandibles with calculus were noted, two of which were described as heavy calculus deposits and the rest being mostly moderate on the lingual side and slight on the buccal side. This is an overall prevalence of 2.0%, which varies from 0.6% in Period 3 to 4% in Period 4-5. This is a very low frequency, particularly when compared to the 24% quoted for Chichester (Levitan 1989). A single mandible showed evidence of periodontal disease, with moderate alveolar recession around the P4 and M1. Such a low prevalence of non-congenital oral pathology suggests that the cattle population was in a generally healthy state throughout all periods of occupation at Heybridge.



Cranial perforations were noted on two fragments, a 5mm hole just below the nuchal suture on one fragment (context 20034, Group 713, Period 4-5) and two irregular holes near the suture on the other (context 14939, Group 713, Period 4-5). The aetiology of this phenomenon is not resolved, although many theories have been put forward (Brothwell et al. 1996). A third cranial fragment (context 14558, Group 722, Period 6) had two holes, one either side of the midline of the facial part of the frontal bone and another hole into the brain case (Figures 605 and 606). These holes were very similar in appearance to cranial perforations but are not in the usual places. Although it is becoming more evident that it may be congenital, as aurochs and wild bison skulls and pigs have been found with these perforations (Manaseryan et al. 1999; Fabis and Thomas 2011), the condition may be exacerbated by yoking to the back of the skull. This would fit with the hypothesis that some of the Heybridge cattle were draught animals (see below). Two horncores displayed 'thumbprints' (Pölloth 1959); one (context 8505, Period 5) was an elongated oval on the posterior aspect near the base of the horncore, the other case (context 4015, Group 739, Period 4) formed a ring-like depression around the base (Figure 607), very similar to examples shown in Müller (1992) and interpreted as an effect of yoking. This condition is more commonly observed in sheep (Albarella 1995).

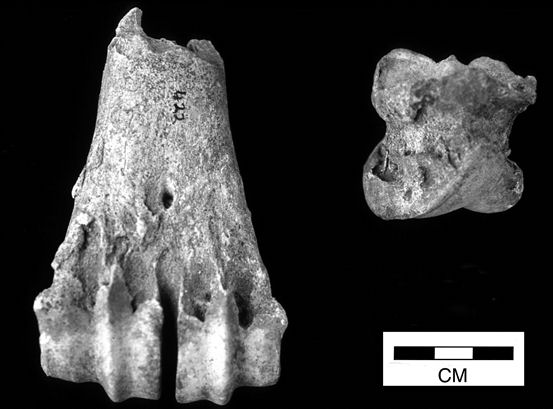

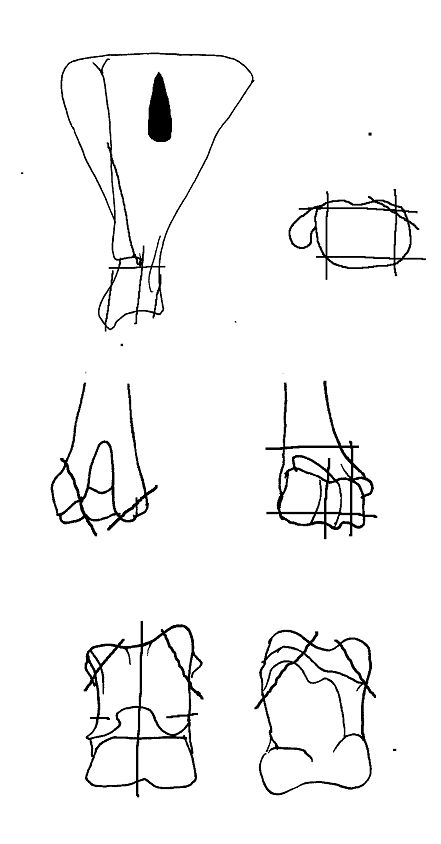
Most cattle pathologies affected the lower extremities of the limbs - distal metapodials and phalanges (Figures 608 and 609). A total of twenty-eight 1st phalanges were affected, displaying splayed proximal articular surfaces (an extension of the articular facet) and/or marginal arthritic reactions around one or more articulations but no signs of degeneration to the articular surfaces themselves. In addition, sixteen 2nd phalanges and a single 3rd phalanx were affected by this kind of pathology. Most only showed slight to moderate splaying and slight marginal reactions but five were severely affected. For the metapodials, only extensively splayed cases were noted, comprising four metacarpals and three metatarsals. One of the metatarsals showed joint surface degeneration (eburnation and grooving) as well as marginal reactions. In addition, four metapodials were noted as being asymmetrical at the distal ends. This showed as a marked difference in the lengths of the condyles (Figures 610 and 611).
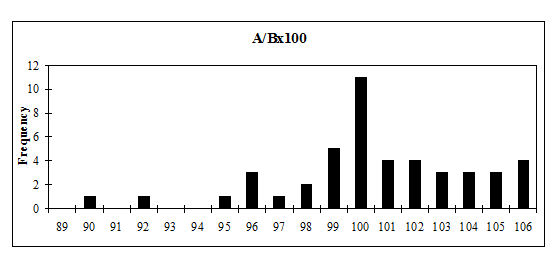
Several measurements were taken on the distal metapodials in an effort to quantify the degree and nature of the splaying observed. This proved to be a very complex problem with many contributing factors acting together. Figures 613-618 show some of the ways the data were used to try to elucidate the problem. Starting with Figure 613, measurements A and B (widths of individual condyles) were expressed as a percentage of each other in order to establish if there was any asymmetry between the two condyles. A value of less than 100 shows that A is smaller than B, and vice versa for values over 100. There are three things to note from the figure - firstly that the modal class sits nicely on 100, showing that 'the norm' is for the two condyles to be of equal width. Secondly, that most values fall within 3% either side of 100, and lastly that there are only two values that appear distinctly asymmetric. This suggests that in most cases the splaying is affecting the condyles almost equally rather than asymmetrically.
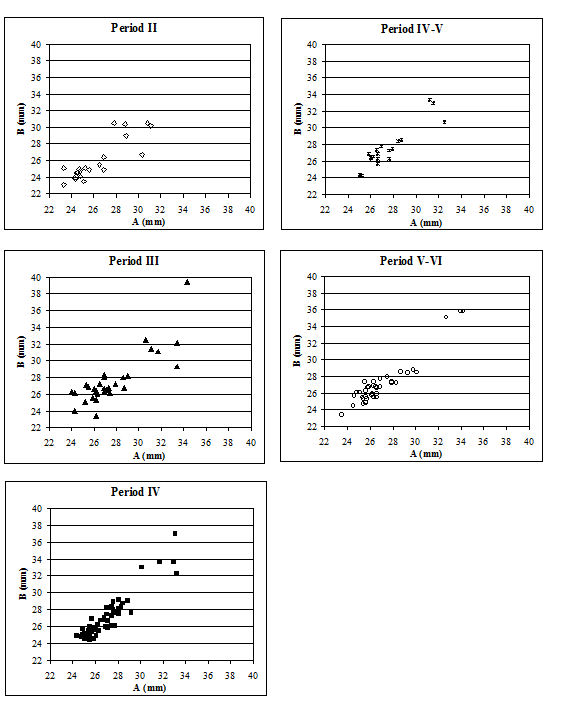
Figure 614 shows the relationship between A and B in metacarpals but as individual points so that bones on which both condyles are splayed may be picked up. In all periods there are a few specimens that form a larger, generally distinct, cluster. To check if these were large or splayed individuals, the specimens giving the large measurements were checked visually. In all cases the larger measurements appear to derive from splayed specimens. This shows that there were many more splayed metacarpals present in the assemblage than were recorded as such in the pathology notes.
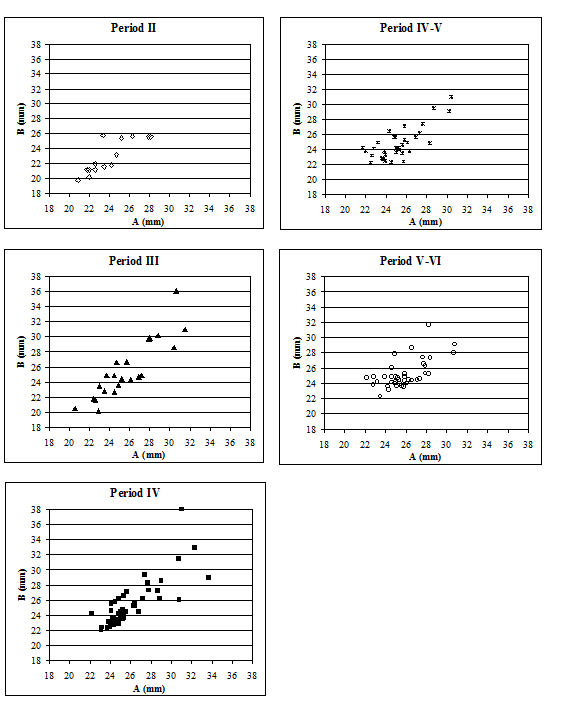
When the same data are plotted for the metatarsals (Figure 615), a slightly different picture is noted. A separation of a group of larger individuals only occurs in Periods 3 and 4, and even in these periods it is not as clear as for the metacarpals. In addition, the points do not cluster as tightly around a linear relationship as the metacarpal measurements. This may suggest that there is more asymmetry in the metatarsals than metacarpals. When the specimens giving larger measurements were checked visually, most did not appear to be splayed and those that showed some splaying were mostly asymmetric. This means that the distal metatarsals at Heybridge provide a better measure of the size of the cattle than the metacarpals, by showing that the outliers are mostly genuinely larger rather than pathological individuals.
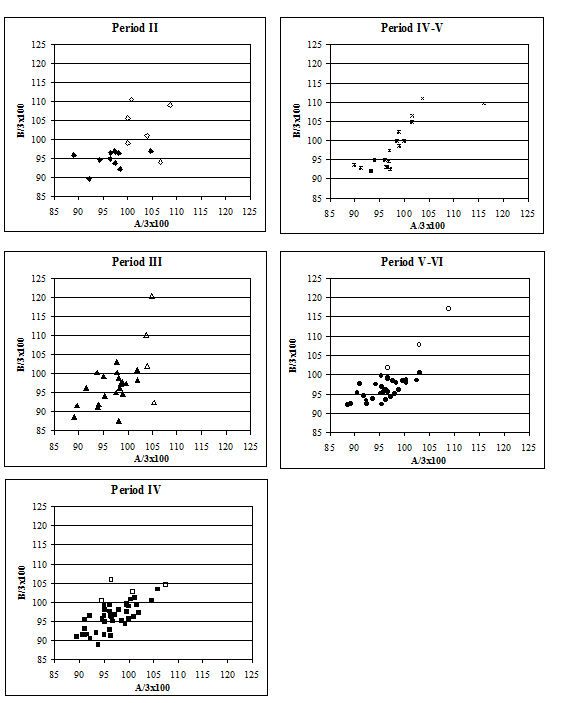
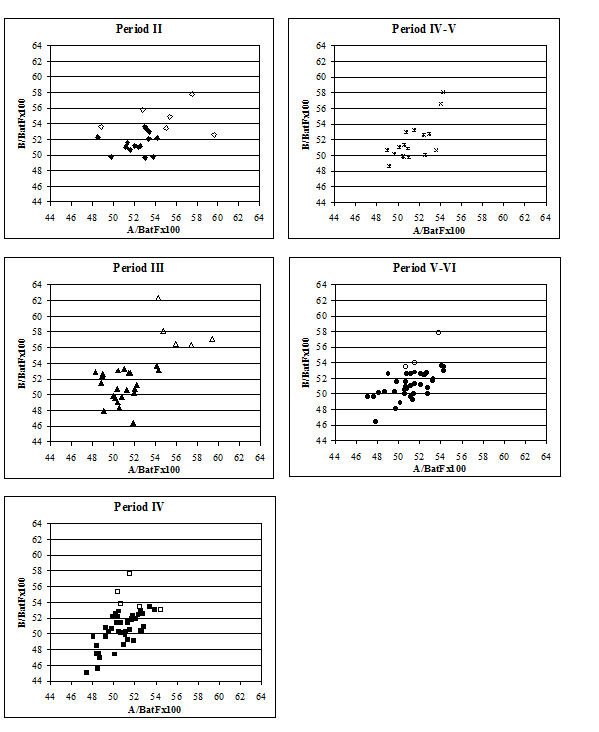
In an attempt to eliminate the confusing factor of the different size of individuals in the analysis of the splaying, diagrams based on measurement indices - therefore size-independent - were calculated. Figure 616 shows the data when the 3 measurement (depth) is used as the denominator. In these diagrams, points located towards the right and the top of the distribution do not indicate large individuals but rather specimens that have condyles that are unusually large in comparison with depth of the distal end. The open symbols were used to distinguish the specimens whose measurements were deemed as 'large' in Figure 614. In most cases the 'large' individuals plot towards the top and the right of the distribution, which confirm what had been visually observed, namely that they are large because splayed. Using another index, with BatF (width at the fusion point) as the denominator (Figure 617), shows a slightly different pattern to Figure 616 but still the 'large' individuals pull away from the main group in most periods.
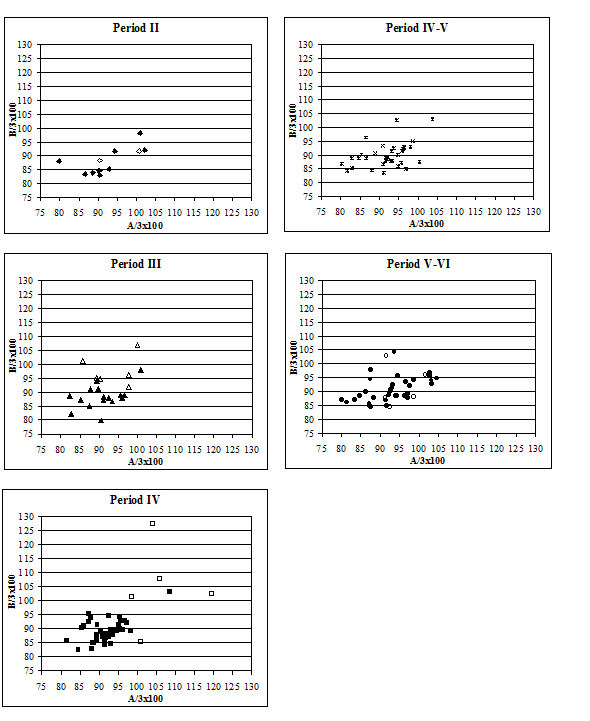
The index of measurements A and B to three on the metatarsal (Figure 618) confirms that many of the 'large' individuals are indeed just large rather than abnormal. In most periods the 'large' group fall well within the main group, suggesting that their shape is normal. Only in Period 4 do most of those with large dimensions also show different shape indices. As with the metacarpals, a few other individuals fall outside the main group in terms of shape suggesting that these may be abnormal, smaller individuals. In summary, there appears to be no one way of looking for splayed metapodials using biometry. A combination of different analyses gives some indication of their presence but no straightforward quantification was possible.
Therefore a visual inspection and classification using the system described by Bartosiewicz et al. (1997) in their extensive study of draught cattle seems to be the best approach. Their classification of the severity of the different types of changes is on a scale from 1 to 4 with 1 being normal and 4 for extreme cases. The values are attributed according to the severity of a number of changes to the bone, including the degree of eburnation and the quantity of marginal bone growth. The scale is applied to both articular ends of the metapodials and phalanges. Most of the cases from Heybridge were either 2 or 3 in severity with only a very few reaching stage 4. It is interesting to note that none of the draught cattle used in the study showed degeneration of the joint surfaces as was the case at Heybridge. The age of the cattle at Heybridge, combined with the relative prevalence of this type of pathological condition of the lower limbs, suggests that at least some of these beasts may have been draught cattle. Although there were too few clear cases for any trends through time to be significant, it does appear that most examples came from post-conquest deposits.
Four types of non-metric traits of the mandible/teeth were recorded. These were the absence of the second premolar, absence or reduction of the hypoconulid (3rd cusp) of the 3rd molar, the presence of premolar foramina and abnormalities of the mental foramen. The third of these traits was only noted in caprine mandibles so will be discussed below. The first two of these traits have been recorded by many zooarchaeologists and the data are now beginning to be better understood.
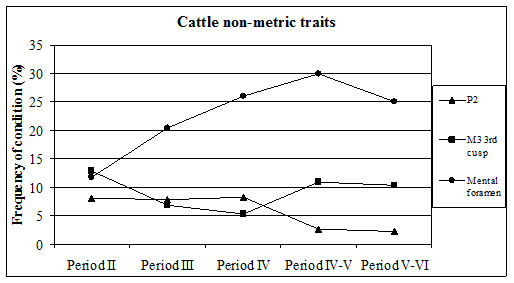
For Heybridge, there are marked changes in the frequency of all recorded traits over time (Figure 612). The absence of the 2nd premolar remained constant at around 8% for the first three periods before dropping to around 3% for the last two periods. In other Late Iron Age sites the numbers of cases are described as rare or infrequent, so Heybridge appears to have a relatively high prevalence in this period. Figures were more readily available for the Roman period and show quite a variation ranging from 4% (The Park, Lincoln; Scott 1985) to 29% (Stonea, Cambridgeshire; Stallibrass 1996). At Chichester (Levitan 1989) there is an increase through time from 11% in the Early Roman period to 25% in the very late Roman period. At Lincoln (Dobney et al. c1996) there is also a rise in prevalence from 6% (1st-2nd century AD) to 12% (3rd-4th century AD). At York (O'Connor 1988) the Early to mid-Roman figure is 19%. It appears that during the Roman period at Heybridge the trend of the prevalence of absent P2s is almost opposite to those elsewhere and is certainly lower in the latest periods than other sites for which figures are given.
The absence or reduction of the hypoconulid was noted on almost 13% of mandibles in Period 2 which falls to 7% in Period 3 and 5% in Period 4 before rising back to around 11% for the final two periods. A few figures were available for this trait in the Iron Age, indicating a range from 7 to 30% although the number of mandibles from some sites was quite small. The Heybridge material at this period fits within the given range. Prevalence in the Roman period shows a wide range, from 2% (Caister-on-Sea; Harman 1993) to 29% (Brancaster; Jones 1985). At Lincoln (Dobney et al. c1996) there is a fall from 10% in the 1st-2nd century to 2% in the later period. At Exeter (Maltby 1979) the prevalence is given as 21%. From these data it seems that Heybridge has quite a low prevalence of this trait in all the Roman periods and that the trend is the reverse of that seen at Lincoln.
The last trait recorded on cattle mandibles was that of abnormally shaped mental foramina. This trait has not been widely recorded but a few examples exist. The abnormalities range from enlarged or elongated single foramina to examples with two completely separate foramina. Photographs of examples are shown in the Lincoln volume (Dobney et al. c1996) and specimens of most types of abnormality were recorded at Heybridge in all periods. The frequency ranged from 12% in Period 2 up to 30% in Period 4-5 and back to 25% in Period 5-6. This is a much higher frequency than for the other traits in the Roman periods. Abnormalities were noted at Lincoln and also at Chichester (Levitan 1989) but the frequencies are not given.
The three recorded non-metric traits seem to bear no relation to each other in terms of their frequency. This is not surprising, as there is no reason why they should be related. The anomalous trends in absent P2 and hypoconulid frequencies in comparison with other sites can be explained by the fact that we still know very little about the variation in this condition. It is, however, possible that the pattern observed at Heybridge reflects the genetic character of the local animal population. Had the population been small and isolated, the gene pool would also be small, which can cause the trait to be over or under-represented.

As a result of the volume of cattle bones recovered from Elms Farm, a large body of biometric data was collected. Summary statistics for all measurements with more than ten cases are given in Table 132 and the individual measurements in the archive. Single measurements were analysed first to study the relative sizes of the bones and teeth between the different periods, before moving on to look at shape indices, log ratios and comparing the results with other sites of the same date.
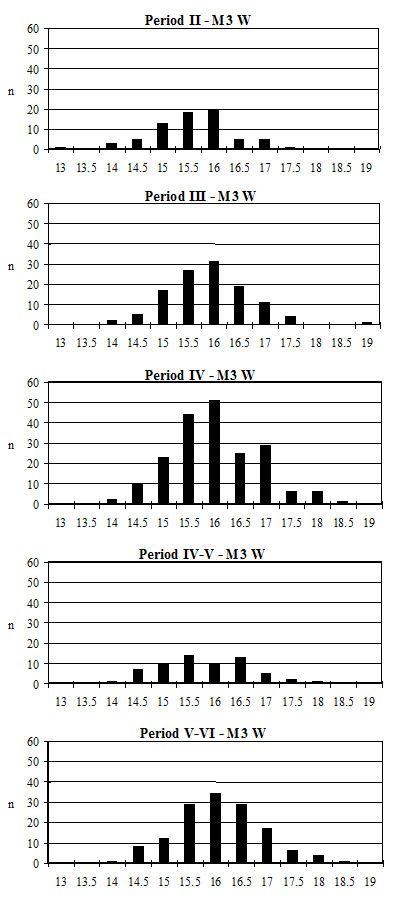
Starting with the teeth, the most frequent measurement was the width of the lower third molar. Histograms of this measurement by period are given in Figure 619. All periods show a reasonably normal distribution centred around 15.5-16mm. There are only slight visible changes between the periods; however, using t-tests to look at this statistically there is a significant increase between Periods 2 and 3 and also between Periods 4-5 and 5-6 (Table 133). Teeth respond very slowly to changes in the size of animals through artificial selection and improved nutrition, unlike the post-cranial skeleton. The fact that the teeth show an increase over time, however slight, should therefore indicate the introduction or evolution of genuinely different genotypes.
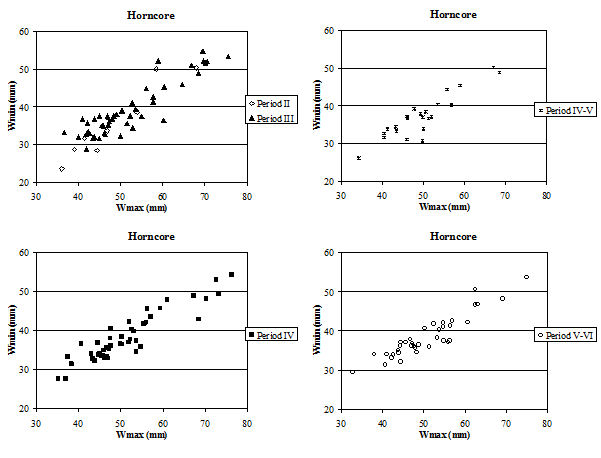
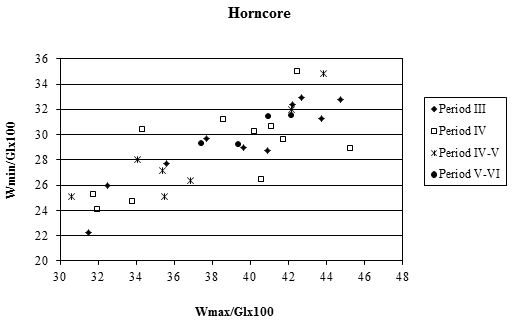
Although there were few complete horncores, many fragments with a complete basal circumference were recovered, which has enabled some analysis of their measurements to be undertaken. Horncores are very variable, both in shape and size, particularly in different sexes and breeds. Figure 620 shows scatter plots of the maximum and minimum diameters of the base of the horncores. In all periods the relationship between the two is roughly linear, indicating that the horncores are all roughly the same shape at the base. The scatter of points in all periods shows a large cluster of points towards the bottom left of the graph and a smaller group towards the top right corner. Figure 621 gives the indices for basal diameter over length for the Roman material, as there were no complete horncores from Period 2. The scatter of points is more open (less linear) than in Figure 620, indicating that there is more variety in shape when the length is taken into account.
There is no clear pattern of difference, and in all periods there seems to have been mostly a rather short-horned type of cattle (Armitage and Clutton-Brock 1976). In all diagrams the specimens tend to group into two clusters. It is possible that the large groups at the bottom of Figure 620 represent females. Castrates would have longer horns with a relatively smaller basal circumference, and might form the small clusters at the top of the diagrams in Figure 620 and at the bottom of Figure 621. These diagrams may not include any entire males, as most of them would be castrated when young and only a few left entire for reproduction. If any were present these would plot in the upper right areas of the scatters in both figures, as bulls tend to have short and robust horns.
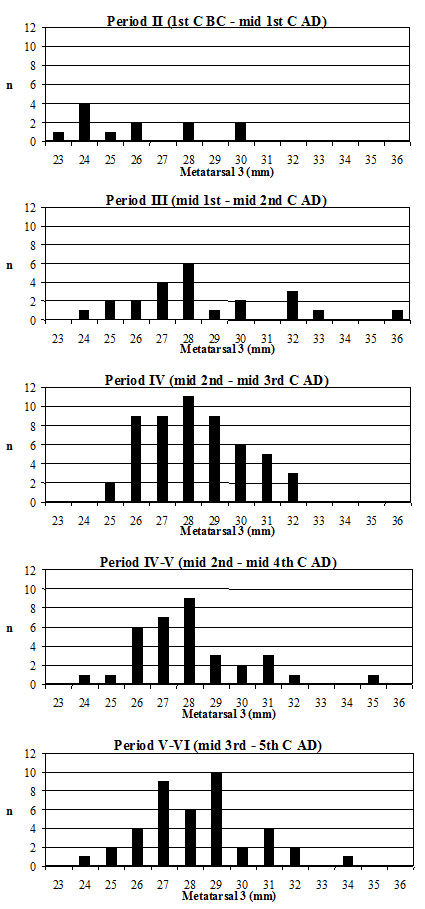
Another commonly used method of looking at the problem of sexual dimorphism in cattle is to use the metapodial measurements. Unfortunately, there are two reasons why this was not possible for the Elms Farm material. The first is that very few complete metapodials were recovered and hence very few greatest length measurements could be taken. Secondly, the distal metacarpals in particular were often affected by pathological splaying, meaning that the measurements of this area may be reflecting this condition rather than the true size of the animals, as discussed above. One measurement was deemed to be of use in looking at the general size of the cattle, measurement 3 on the metatarsal, as it was least likely to be affected by splaying, being a depth and therefore located on a different axis. Figure 622 show histograms of this measurement by period. Although there are not many data for Period 2 these are significantly smaller than the Period 3 material and highly significantly smaller than the later periods (Table 133), while there is no significant difference between the later periods. This indicates that the size of the cattle increased at around the time of the Roman conquest, suggesting that the invading army may have brought sufficient numbers of larger cattle with them to improve the local stock almost immediately.
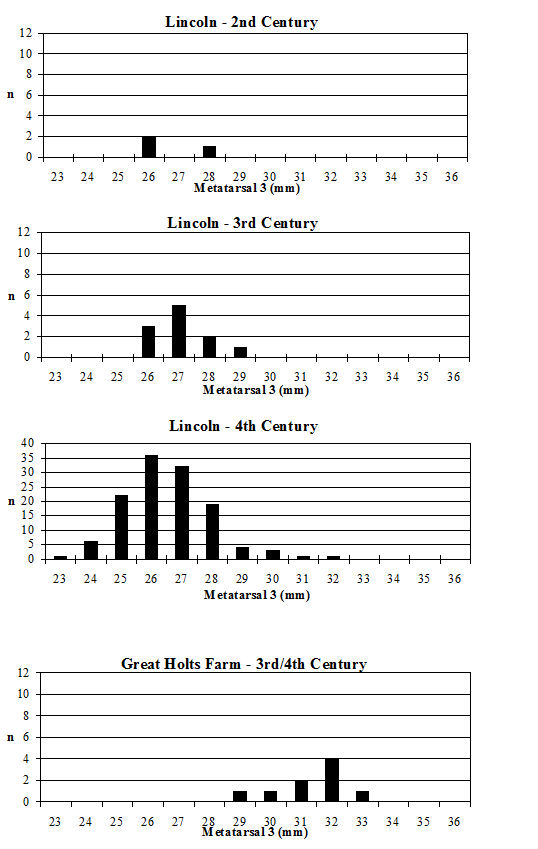
Looking at the Heybridge data in a wider context, the Heybridge material is considerably smaller (t-test highly significant) than the cattle bone from Great Holts Farm (3rd/4th century AD) (Albarella 1997) in the comparable periods (4-5 and 5-6) (Figure 623). However, in comparison to material from Lincoln at the same period (Figure 623, 4th century) the Heybridge material is notably larger (t-test highly significant). Although there is very little data from the 3rd century in Lincoln, this is significantly smaller than the Period 4 material at Heybridge. This suggests that although at Lincoln there was a decline in the size of cattle in the 4th century (Dobney et al. c1996), such a size decrease did not occur at Heybridge. The very large cattle at Great Holts Farm are certainly exceptionally large, and Albarella (1997) concludes that they may represent first-generation imported breeding stock (the site is a villa). Following from this hypothesis, the cattle at Heybridge may be slightly smaller as a result of cross-breeding.
The size of the astragalus is directly related to its weight-bearing role and is therefore also closely related to the size of the animal. This makes it a particularly good bone to use in biometric studies as a measure of the robustness of the individuals. Figure 624 shows histograms of the Bd measurement by period. The same picture emerges from this measurement as from the metatarsal (Figure 622), with a highly significant increase in size occurring between Periods 2 and 3. Bd is a measure of the width of the joint surface and is therefore related to the weight-bearing ability of that joint. This suggests that an increase in this measurement reveals the presence of significantly more robust individuals rather than those that are just taller. Comparing the animals from Lincoln (Figure 625) with those from Heybridge, there are some similarities but more differences. The 1st-century Lincoln material appears to be smaller than the Period 3 material but there are too few cases to test the significance of his. The 3rd-century material appears to be similar to Periods 3 and 4 but again there are too few cases to test. The 4th-century material appears smaller than the latest two periods at Heybridge and this difference is highly statistically significant (Table 133).
Figure 626 gives the shape indices for the cattle astragali and show no changes between the periods. They also show that there is no linear relationship between the two indices so there is quite a lot of variation in the shape of the astragalus. There are however, no clear groupings, which would indicate that most animals, even in different periods, were of similar builds. This is interesting as it might be expected that different sexes, and particularly castrates, would be slightly different. Perhaps the differences are too slight to stand out visually.
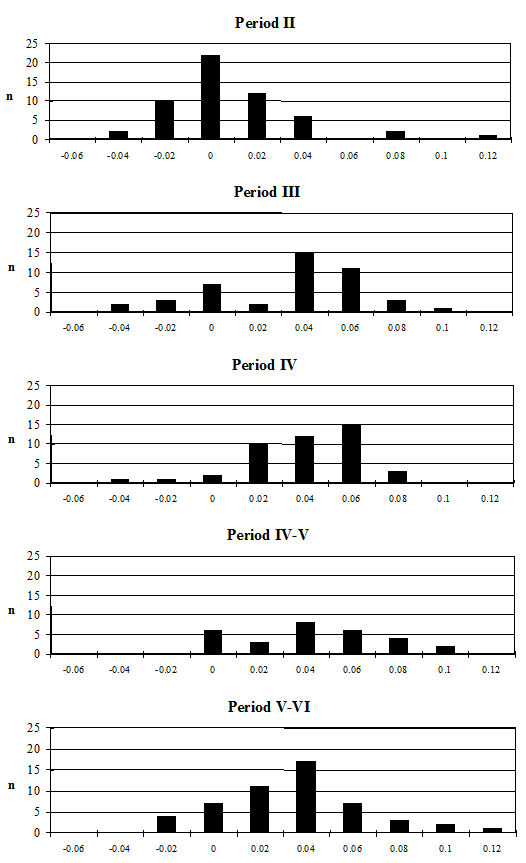
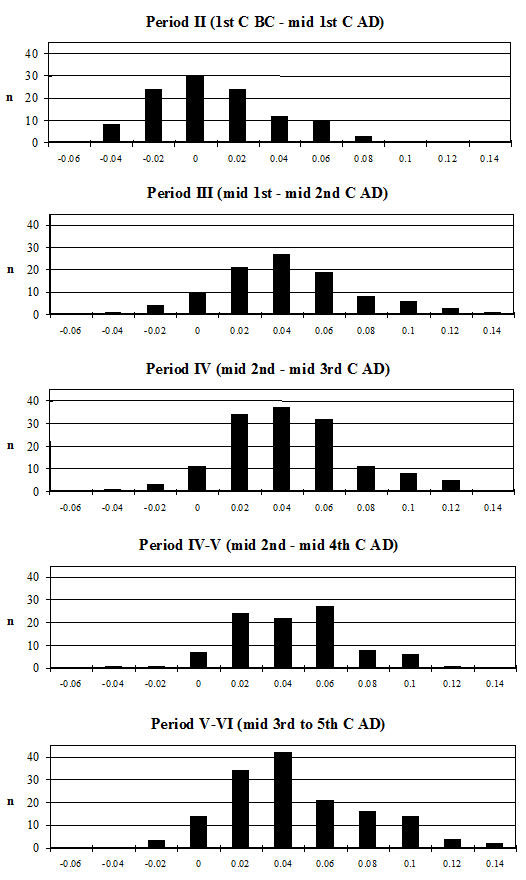
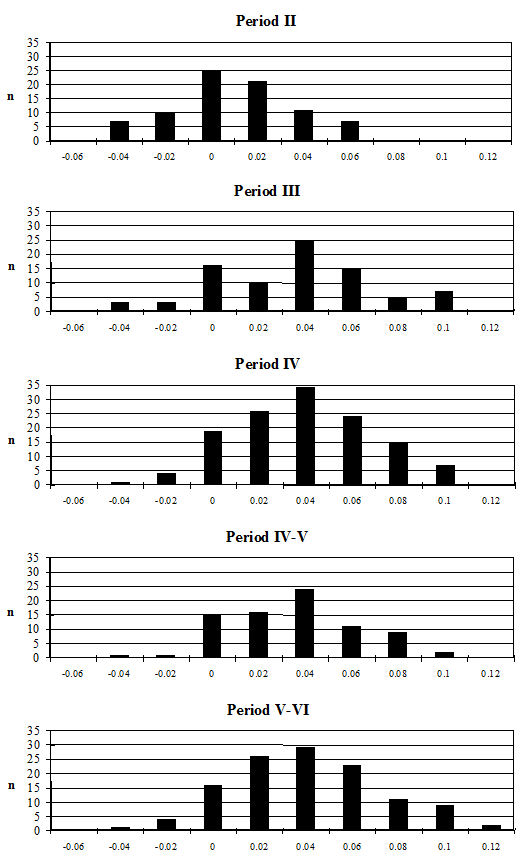
The remaining measurements all had too few cases to be analysed individually or were unsuitable for other reasons (such as pathology). In order to maximise the information potential of the data, log ratios were calculated to allow different measurements to be combined. As suggested by Davis (1996) the length, width and depth measurements were kept separate, as there is better correlation between measurements taken on the same axis than between those on different axes. As there is no published British standard to calculate cattle log ratios, the means of the data from Period 2 were used. When the data are plotted, Period 2 will always be centred on zero giving an easy reference point to indicate whether material from other periods is larger or smaller than Period 2. Figures 627, 628 and 629 show the log ratio diagrams for the lengths, widths and depths respectively. Once again there is an increase in size between Periods 2 and 3. In all three dimensions this increase is statistically highly significant (Table 133) while there is no difference between the later periods.
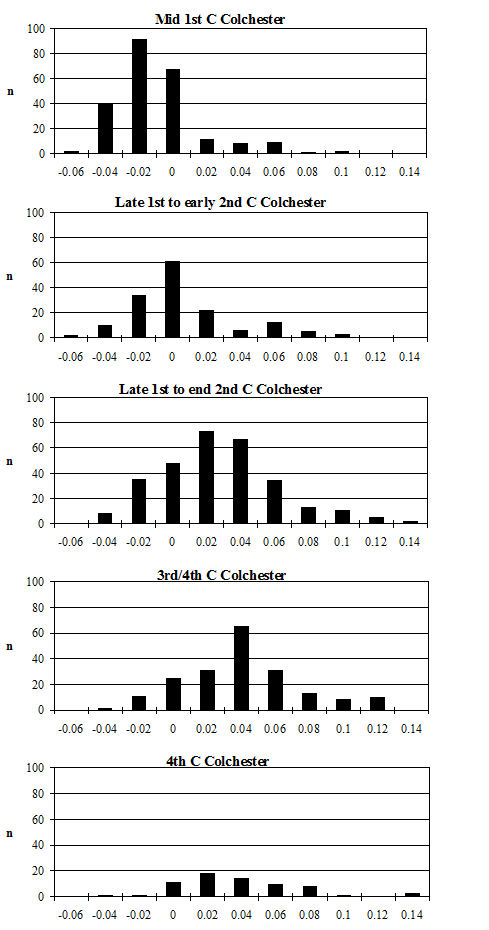
For comparison with other sites, only the width measurements have been used as these were generally the most numerous. The pattern at Colchester (Figure 630) is somewhat different from that at Heybridge despite their relatively close proximity. There is again a size increase but it happens at a slightly different time and to a lesser degree. Within the Colchester assemblage itself there are highly significant increases in size between the first four graphs in Figure 630. Only between the 3rd/4th and 4th-century material is there no change. This indicates a steady change in size through time rather than a sudden increase that is subsequently maintained. When compared to Heybridge the earliest material (mid-1st century) from Colchester is very significantly smaller than the Period 2 material at Heybridge, which suggests that even prior to the conquest the Heybridge material was relatively large. The next two periods at Colchester are also significantly smaller than their corresponding periods at Heybridge (Table 133) but the 3rd/4th-century material is no different from the Period 4-5 and 5-6 material at Heybridge.
The pattern at Colchester may well reflect the fact that this is a large urban centre which, being a predominantly consumer rather than producer site, is getting its animals from a far greater geographical area than Heybridge (probably a producer and consumer site). This means that animals with a far greater size range will have become part of the bone assemblage, many of which may not have been as 'improved' as the Heybridge stock. This hypothesis is supported by the fact that, despite the average size of the Colchester cattle being smaller, a few animals are as large as the largest ones found at Elms Farm. Eventually the animals at Colchester did reach the same stature as the Heybridge ones as the process of livestock improvement continued.
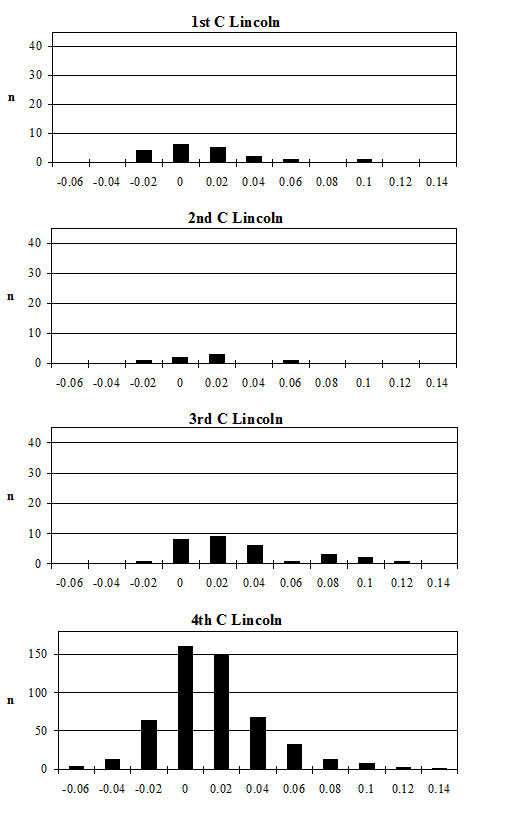
Looking further afield, the data from Lincoln are presented in Figure 631. Again the picture differs slightly, both from Heybridge and Colchester. There are very little data for the 1st and 2nd centuries at Lincoln but cattle size is similar to Period 2 at Heybridge and to the same periods at Colchester. There is no marked size change between any of the periods; a slight increase may be apparent between the 1st and 2nd centuries and there is a significant increase between the combined 1st and 2nd century data and the 3rd century. There is also a significant decrease from the 3rd century to the 4th century. The 3rd-century data from Lincoln is not significantly different from Period 4-5 at Heybridge but the 4th-century data is very significantly smaller than the Period 5-6 data. This phenomenon of a decrease in the size of the cattle at Lincoln in the 4th century has been noted for other measurements (discussed above) and is confirmed here, as is the fact that no decrease occurs at Heybridge.
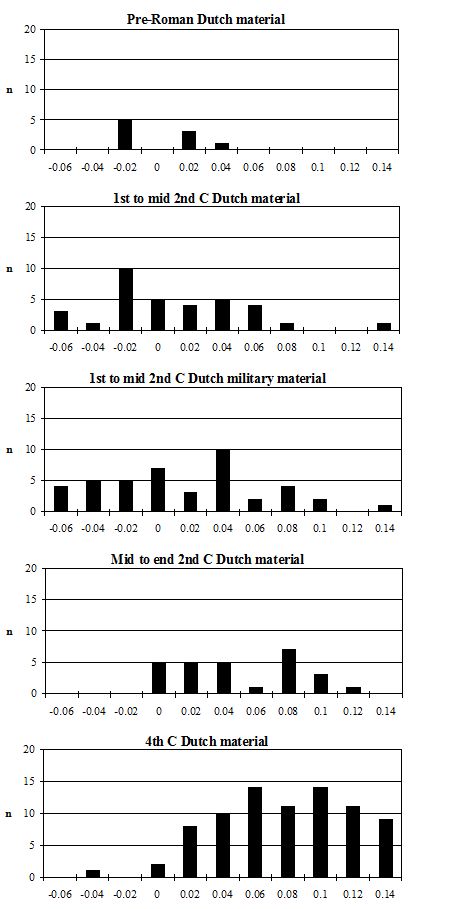
Comparing Heybridge with data from the European mainland (Figure 632), yet another picture emerges. Both Dutch civilian and military data sets show that the cattle were highly significantly smaller than those from Period 3 at Heybridge. There is, however, a size increase in the Dutch cattle from the middle to end of the 2nd century, which are significantly larger than both the preceding civilian and military assemblages. In the 4th-century material there is a further significant increase. There is no significant difference between the mid-end 2nd century Dutch material and Period 4 at Heybridge but the 4th-century material is very significantly larger than the later periods at Heybridge. The Dutch material appears to be slightly smaller than Heybridge just after the conquest of Britain, but increases in size from then, overtaking the Heybridge animals in the 4th century. Perhaps this suggests that while the largest animals were imported to Britain to improve the native stock immediately, the improvements did not continue in later periods as seems to have happened in Holland. Alternatively, and perhaps more likely, the progressive increase in size that we witness for the Netherlands did not occur in Britain as a consequence of a drought in the importation of new stock by the Late Roman period. This would be consistent with a general decrease in the intensity of overseas trade that is known for 3rd- and 4th-century Britain (Fulford 1989; Millett 1990).
Taken all together, the cattle biometric data presents a relatively coherent picture. There was a deliberate and substantial improvement in the size of stock in the immediate post-conquest period, which was sustained but not improved further throughout the occupation of the site. The immediacy of the improvements suggests that Heybridge was close to (or was itself) a producer site while on larger urban consumer centres the increase is not seen until later. The lack of evidence for a decline in size in the 4th century, as seen at Lincoln, perhaps suggests that the area around Heybridge was more highly Romanised and continued to use Roman practices of stock management, even when the rule of Rome was beginning to fade in the northern provinces.
The identification of sheep and goat bones to species level was attempted on a limited range of elements (see methods), the vast majority of which were identified as sheep (Tables 118 and 119). Only two definite, and three tentative, goat identifications were made in the whole assemblage. Such a low incidence of goat bones is a common occurrence on British sites of all periods.
The goat bones were spread across all periods, which suggests that they were present in small numbers in or around the settlement, throughout its history. Goats can thrive on rough pasture and human domestic refuse and are therefore an ideal animal, together with pigs and chickens, to keep in a confined area such as a back yard. Although sometimes eaten, goats were primarily kept for their milk, as the yield was higher and the products more desirable than ewes or cows milk (White 1970).
As such a low percentage of the identified remains were goats, it has been assumed that most of those recorded as caprine were sheep rather than goats, and the discussion that follows is, therefore, centred on sheep rather than both species.
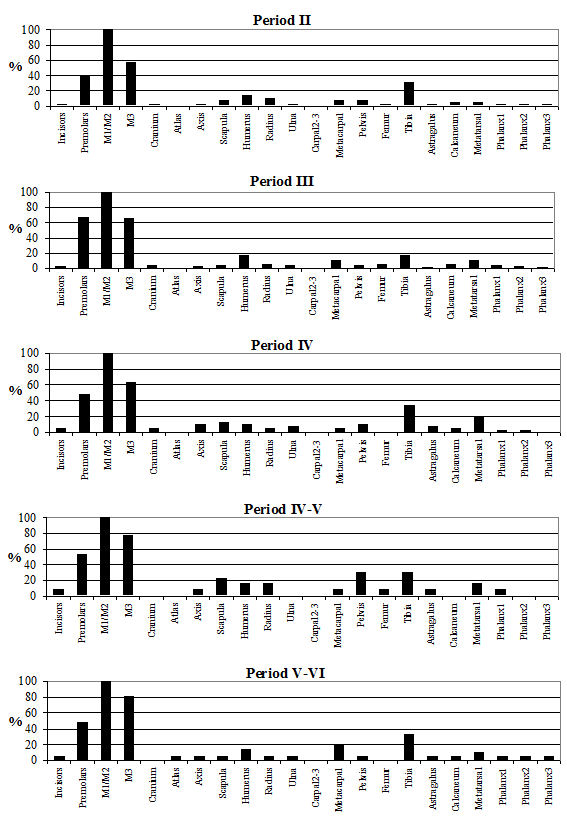
The skeletal element distribution for the sheep/goat bones is given in Table 123 and Figure 633. In all periods the highest MNI value was gained from the M1/M2; this was forty-two in Period 2, eighty-seven in Period 3, thirty-eight in Period 4, thirteen in Period 4-5 and twenty-one in Period 5-6. The distribution patterns were similar to those for cattle, excepting that the tibia was usually the best represented among the post-cranial material. The proportion of teeth to post-cranial material was similarly large, probably again due to the differential preservation of bones and teeth. The small elements were conspicuously absent in the sheep/goat material, as would be expected owing to the inevitable size bias in hand-collected material. An analysis of the distribution of material by context type (Figure 634) and area (Figure 635 .pdf) shows no obvious differences in the proportions of skeletal elements between context types or areas.
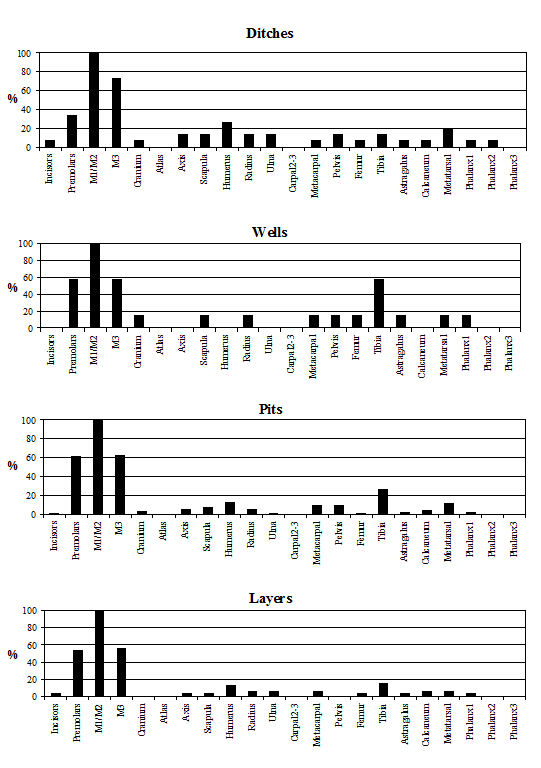
As with cattle, the skeletal element distribution for the caprine material indicates that the animals were likely to have been killed, butchered and consumed on site with no obvious deposition of parts utilised for a specific purpose. This distribution pattern is seen on most sites for which no specific industrial or craft refuse is present and which were not large enough to generate the amount of refuse that would warrant a more organised rubbish disposal procedure.
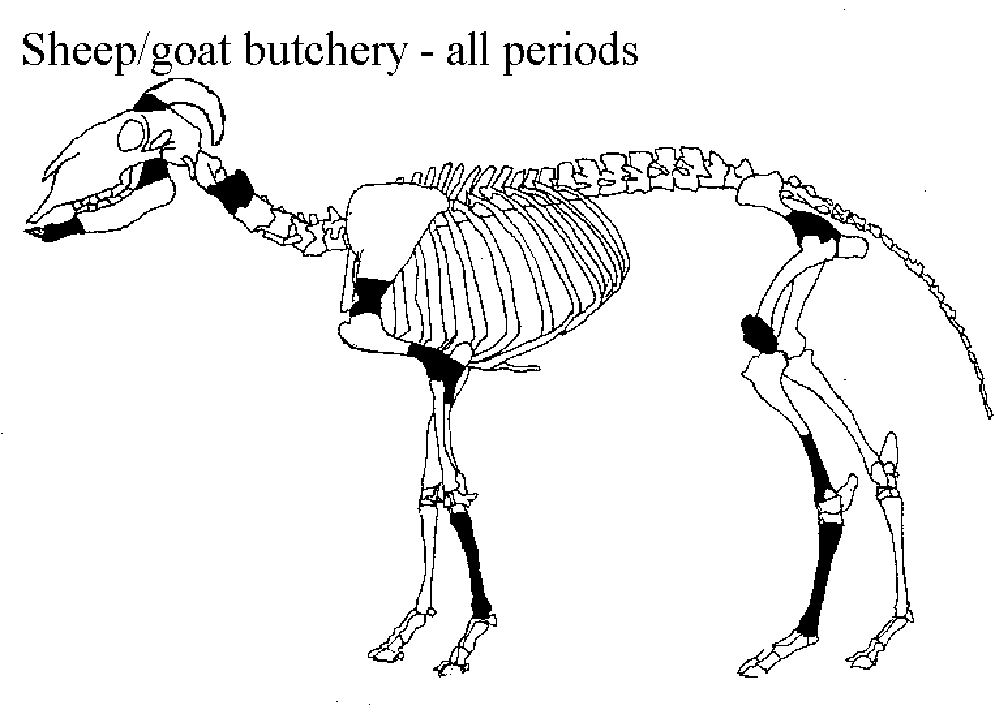
Jointing of carcasses caused some butchery on post-cranial elements, but the frequency of these marks is low (Figure 636). A few longitudinally split metapodials were noted (contexts 9610, Group 75, and 4459, Group 755, Periods 2 and 3 respectively), indicating possible marrow extraction. Several horncores had been separated from the cranium at the base of the horncore (context 13398, Group 409, Period 3 and context 4954, Group 739, Period 4), suggesting that the horns were utilised. A single mandible had been chopped through the diastema (context 4197, Group 744, Period 6) and two through the ramus (contexts 23324, Group 31, Period 2 and 13398, Group 409, Period 3) probably to enable the tongue to be removed from the head. In most archaeological sheep/goat assemblages chopping marks are not abundant, perhaps because sheep/goat carcasses were usually dismembered using a knife to cut through the tendons and joints. This was corroborated by the fact that two metapodials (contexts 4403, Group 741, Period 5) and 5418, Group 422, Period 4)) had horizontal knife marks around the proximal end, suggestive of either skinning and/or jointing at this point.
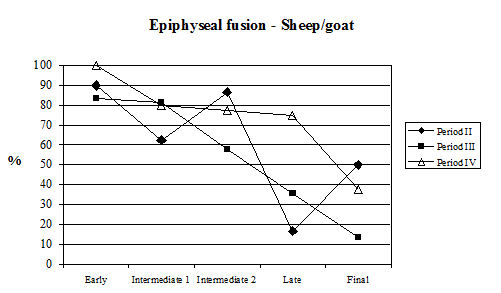
The epiphyseal fusion data for the sheep gives a rather confusing picture, particularly in Period 2 (Table 134, Figure 637). This is probably a reflection of the small sample size in that period. The samples were even smaller in the latest two periods and the results are therefore not plotted. Periods 3 and 4 show slightly different patterns that may be true reflections of the trends in these periods. Period 3 shows a steady decrease in the number of bones fused in each category after Intermediate 1. This indicates that while most of the sheep reached around 18 months of age the number surviving to skeletal maturity was relatively small. In Period 4 there is a sharp fall between the Early and Intermediate 1 groups, there is then a very slight decrease in numbers to the Late group and then another sharp fall to the Final category. This suggests that there were two main age groups being killed off, the first between 10 and 18 months and the second after three years old (using Silver's (1969) data). This suggests that one group comprised lambs killed for their meat and the other group consisted of adult animals killed for mutton but in their prime, rather than after prolonged breeding or use for secondary products.
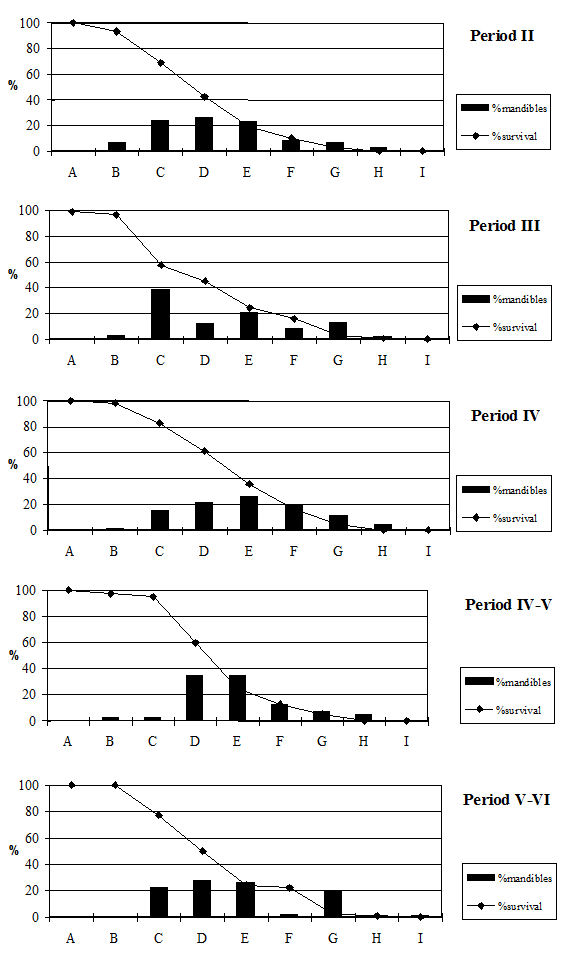
Tooth wear stages were analysed using the categories of Payne (1973) and the results are given in Table 135 and Table 136 (.xlsx) and Figure 638. These show that in all periods two-thirds of the animals had been killed by the time they reached wear stage E at around 2-3 years of age. The kill off patterns were relatively similar between the periods, except Period 3 which stands out as having a larger peak of younger animals (stage C, 6-12 months). The two latest periods have very few mandibles overall, which renders any interpretation of the data unreliable. Periods 2 and 4 are slightly different, with individuals in Period 4 being on average slightly older than those in Period 2 at the time of death.
The two age groups seen in the epiphyseal fusion data do not appear in the wear stage data. It is not unusual for differences of this nature to occur, but, as the bones are more prone to be affected by taphonomic factors, the tooth data probably provide a more reliable picture. With the exception of Period 3, the sheep seem to have been kept for a mixed purpose. Mutton production was probably the main concern but an age of 2-3 or more years old is beyond the point when the maximum body weight is achieved, therefore secondary products, such as wool and milk, were also economically important. Males (probably mainly castrated) were kept until they had produced at least one if not more fleeces, and females were kept until they had lambed at least once, and hence produced milk and offspring as well as the fleeces. The appearance of younger animals in Period 3 may be a response to the growth of settlement at this time and the need to provide food for an increased population.
Lincoln appears to have a similar sheep exploitation pattern (see Dobney et al. c1996), but other Roman sites, such as Exeter (Maltby 1979), show mortality curves more similar to that observed in Period 3 at Heybridge, with most animals being killed towards the end of their first year. Most of the sheep mandibles from Colchester (Luff 1993) were also aged to Stage C; however, on a few sites in specific periods older individuals were noted. Perhaps the differences between sites and dates across the country may be the product of the market economy - when there is a demand for meat, younger individuals are killed to fulfil the demand and when the demand for secondary products is greater the sheep are only eaten as older individuals.
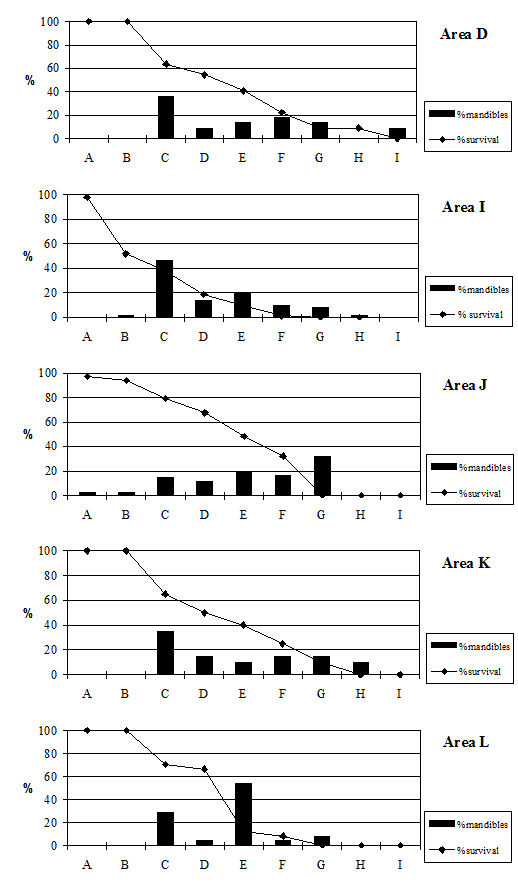
As the pattern observed in Period 3 seemed to be considerably different from the other periods it was decided to look at this material in more detail. The suggestion was put forward that as the temple complex was most active during this period, perhaps this was influencing the kill-off pattern. Figure 639 shows the Period 3 mortality profiles by area and confirms the hypothesis that the temple precinct (Area J) was different in some way, but not in the expected manner. It was expected that the young animals would be associated with the temple complex, as at Harlow (Legge and Dorrington 1985) and Caesaromagus (Luff 1992). However, all the other areas show high numbers of young animals and the temple complex has a high number of older adult animals.
This analysis brings about two main considerations. The first is that the killing of younger animals that typifies Period 3 is found in all areas but one, and this reinforces the hypothesis that such culling strategy is a genuine character of this period. Secondly, if we accept the assumption that the bones in Area J are directly associated with the temple and its activities, it follows that the sacrificed animals would mainly have been represented by mature individuals. Perhaps the reason for this was that young animals were too precious a commodity to sacrifice and that animals past their 'sell-by-date' were used instead.
Examinion of the distribution of mandibles giving adult ages shows that they are all located in two pit complexes, one directly in front of the temple cella to the side of its entrance (Group 409, contexts 5146, 5151, 13398 and 13400) and the other directly behind the temple building (contexts 13200, Group 1137, and 13250, Group 3029), opposite the entrance. Because of the proximity of these pits to the temple buildings it has been suggested that they are probably associated with sacred activities, even though no 'votive deposits' were definitively identified at the time of excavation (Atkinson and Preston 1998). The presence of the mandibles in these pits may give further evidence of their links to the temple complex and its activities in the form of sacrifices. As far as parallels are concerned, the temple complex itself is somewhat unique and the only other Roman site known to the authors where possible adult sheep sacrifices were present is Ivy Chimneys, Witham (Luff 1999).
Of the eight pathological caprine bones, six are represented by cases of oral pathology. An isolated lower M3 (context 5149, Group 3024, Period 3) showed linear hypoplasia across all three cusps around 10mm from the cemento-enamel junction. Two instances of calculus formation were noted, both from Period 3. Periodontal disease (Period 5 and 6), abscess (Period 6) and tooth rotation (Period 5/6) were the other conditions noted.
The two other pathological bones were a sheep horncore (context 14073, Group 276) with a 'thumbprint' manifested as a large shallow depression on the medial side, causing the posterior border to look very pinched. Horncore depressions have been related to nutritional and environmental stress (Albarella 1995), and the fact that these are not common at Heybridge might indicate that sheep flocks were generally in good health. The second was a metatarsal with a spur of new bone formed vertically from the posterior side of the proximal articulation, buttressing the medial posterior ridge for about 20mm. Although this is similar to a condition noted by other workers (O'Connor 1984; Dobney et al. c1996), it is an unusual example as it reaches right to the articular surface and is on the posterior rather than anterior side. The cause of this condition is still unknown.
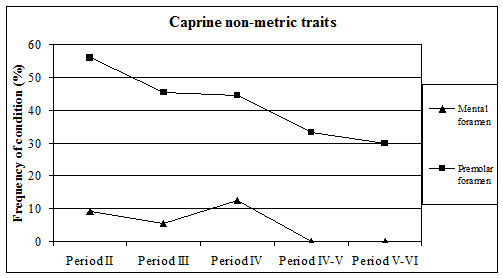
All four non-metric traits described under the cattle pathology section were recorded for the caprine mandibles (Figure 640). However the instances of absent P2 (two cases) and hypoconulids (one example) were so rare that they do not warrant discussion in detail as this level of prevalence appears to be the norm across other sites (O'Connor 1988). Abnormalities of the mental foramen were noted in the first three periods at Heybridge, with percentages of 9.1, 5.5 and 12.5 respectively. As there are no comparative data it is not possible to comment on this trend. The presence of a premolar foramen in the caprine mandibles was very high in all periods and there is a clear decreasing trend from 60% in Period 2 to 30% in Period 5-6. There appear to be two steps in this trend, the first between Periods 2 and 3 and the second after Period 4. This may reflect the introduction of new stock, without the trait to the area, which would 'dilute' the trait within the population. Again, no data are available from other sites to allow comparisons to be made.
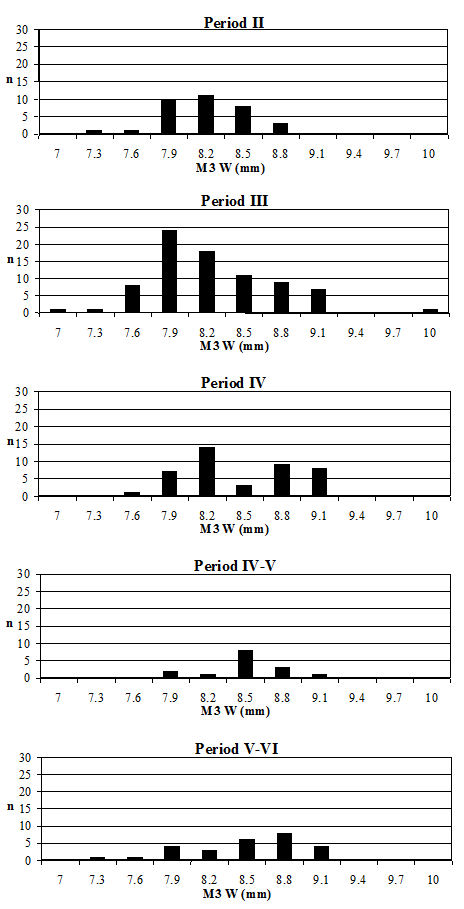
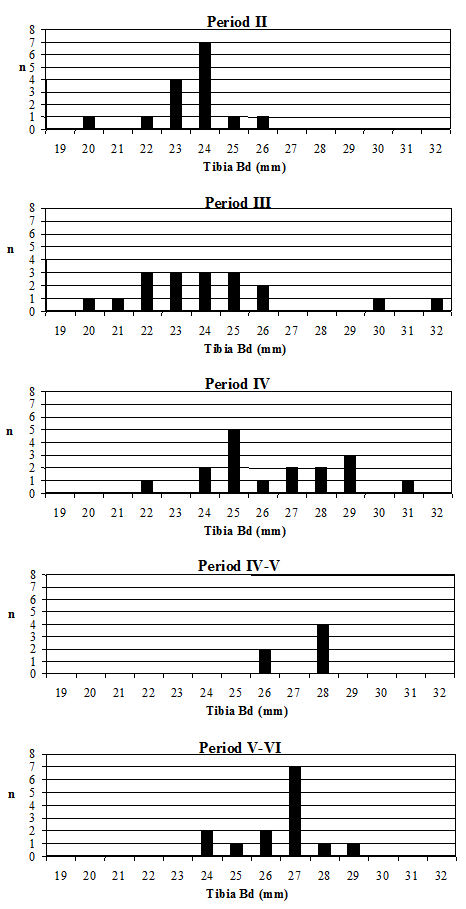
Caprine measurements are not nearly as numerous as those of cattle bones (because of the relative proportions of species), but they can still provide some interesting information. Table 137 gives a summary of the measurements with more than ten cases and individual measurements are given in the archive. The measurements of the teeth were the most numerous, which was expected given the high proportion of teeth in the body part representation. Figure 641 displays the widths of the lower 3rd molar on histograms by period. Visually there is a slight size increase between Periods 3 and 4 which, when statistically tested, gives a significant result (Table 138). The differences between the other periods were not significant. The only post-cranial measurement with enough cases to analyse by period was the Tibia Bd (Figure 642). As with the M3 measurements (above) a significant size increase occurs between Periods 3 and 4 and the difference between Periods 2 and 4 is highly significant. The interesting thing to note in the Period 3 material is the presence of two individuals that are quite a lot bigger than the rest of the material. These could be larger animals (probably male) introduced to improve the local stock. However, the number of cases was too small to test if these were statistical outliers.
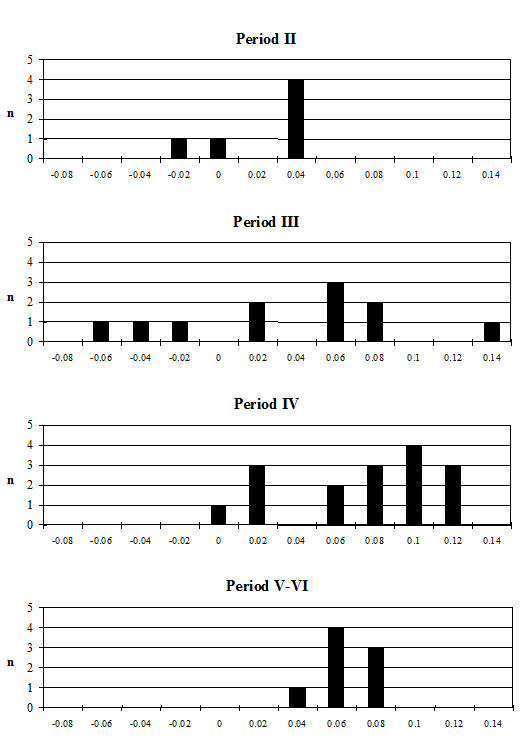
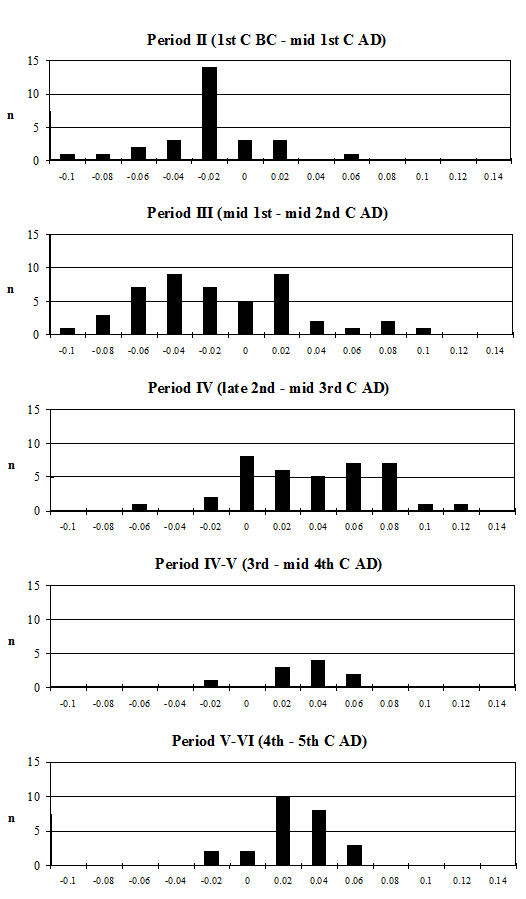
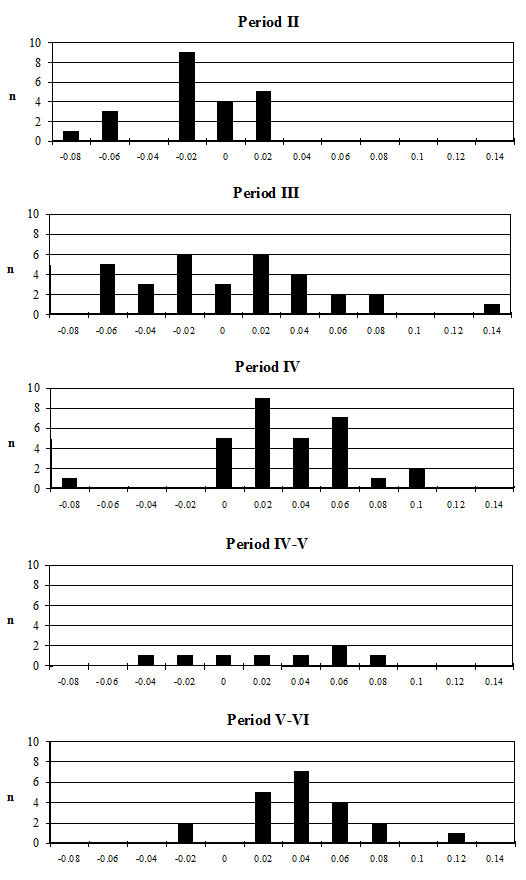
In order to obtain large enough samples to make better comparisons between periods and between this site and others, log ratios were calculated. Only Periods 3 and 4 (Fig 643) provided enough length measurements for a statistical comparison to be made and, as with both measurements looked at so far, the difference between the results of these two periods was significant. Although more measurements were available for both widths (Figure 644) and depths (Figure 645), the pattern that emerges is exactly the same, with the highly significant increase occurring between Periods 3 and 4. The standard used for the log ratio calculations was the mean of a group of Shetland ewes (Davis 1996). In terms of absolute size the Heybridge sheep bones in Periods 2 and 3 were about the same length and depth as the Shetland ewes but were somewhat smaller in the width measurements. For the later periods the Heybridge sheep were slightly larger than the Shetland ewes in all dimensions but again taller in comparison to width and this time to depth also.
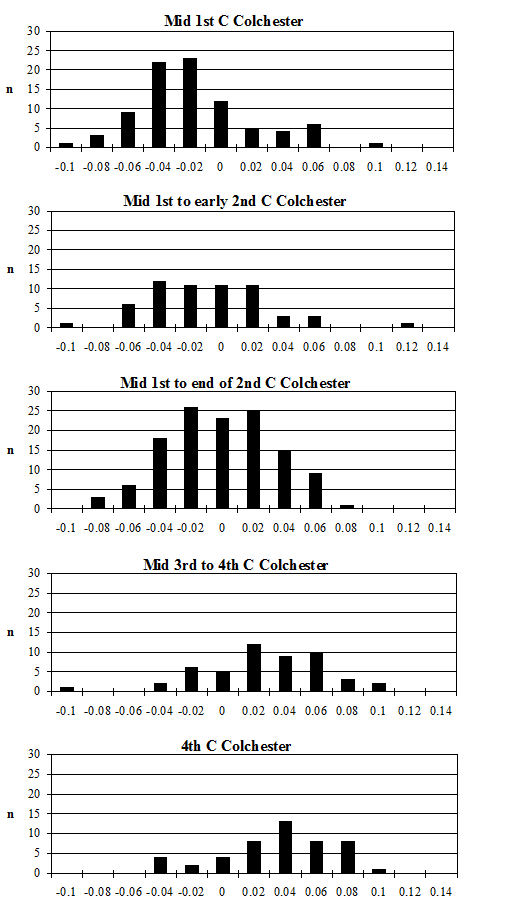
Enough biometric data were available from three sources for comparison with Heybridge; Colchester (Luff 1993), Lincoln (Dobney et al. c1996) and a synthesis of Dutch sites (Lauwerier 1988). Only width measurements were used, as these were most common. There were enough measurements in tightly dated groups for comparisons between the Colchester material (Figure 646) and all periods at Heybridge. The Colchester material showed a very similar pattern to that from Heybridge with an increase in size occurring sometime in the 2nd century AD. The mid-1st to end of 2nd century Colchester material was compared to Periods 3 and 4 individually, and also combined, as it covered parts of both periods. The t-test results (Table 138) showed that the Colchester material was significantly larger than that from Period 3, but smaller than the Heybridge sheep from Period 4. No difference was detected when it was compared to the combined assemblage from the two Heybridge periods. This suggests that the Colchester material in this period is showing the same increase at around the same time but the dating is slightly too broad to pick up exactly when it occurred.
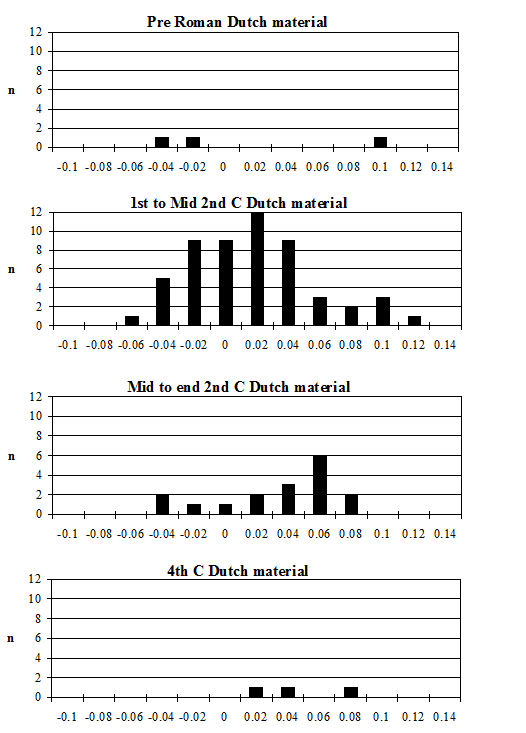
The Lincoln data (Figure 647) were too sparse in the 1st and 2nd century for t-tests to be applied but the size of the sheep appears similar to Heybridge at this time. This is certainly true for the 3rd and 4th-century material where no significant differences between the two sites were observed. At Lincoln too the increase seems to have occurred sometime towards the end of the 2nd century. Looking at the Dutch data (Figure 648) the same increase in size is observed but it happens at an earlier period. The size increase happened before the 1st to mid-2nd century, as that material is significantly larger than the material from the equivalent period at Heybridge (Period 3). Since the Romans conquered Holland a century before Britain, by the end of the 1st century the process of Romanisation had been taking place there already for more than a century. It is therefore not surprising that by this time the improvement in the Dutch sheep was greater than in the British ones.
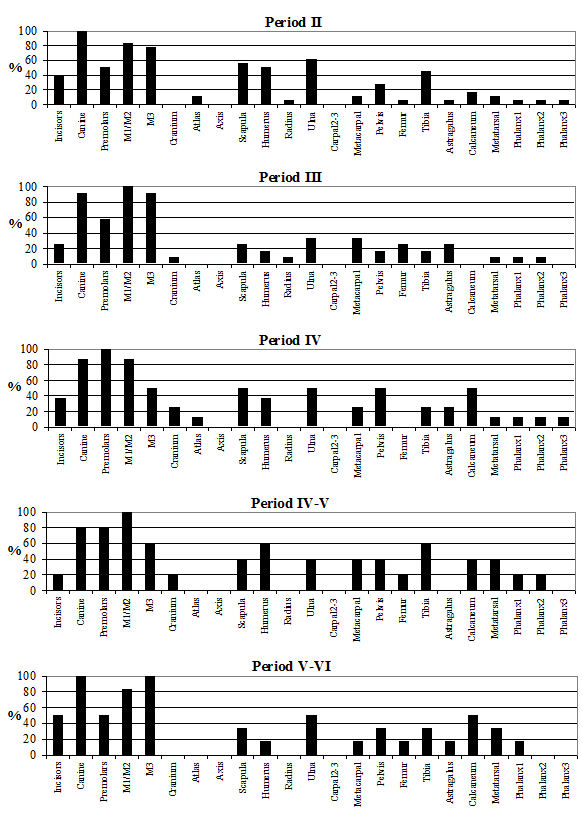
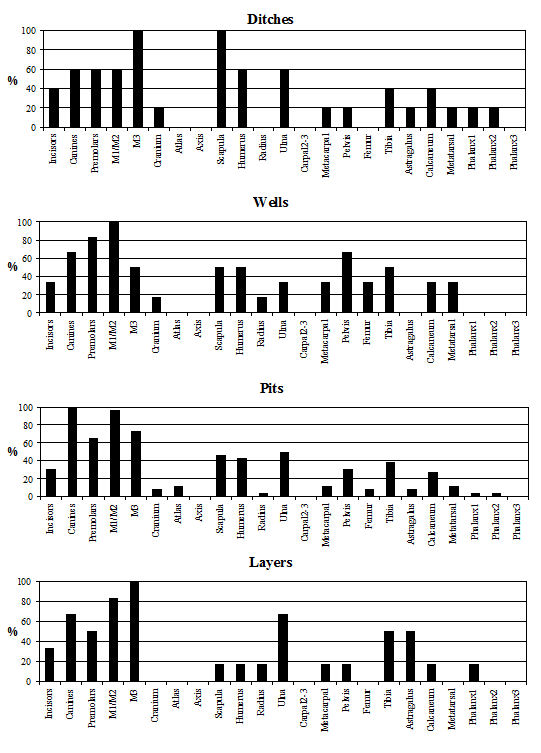
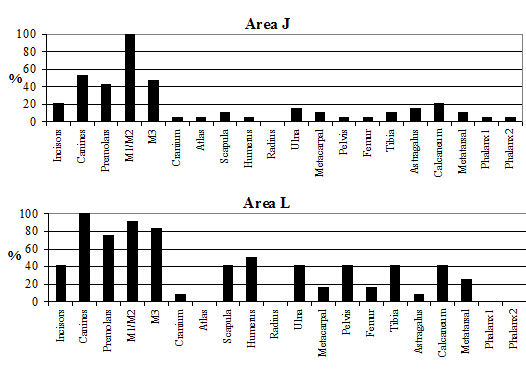
The skeletal element distribution for the pig bones is given in Table 124 and Figure 649. The graphs show the percentage MNI, where the highest values (100% in diagrams) were 18 in Period 2, 12 in Period 3, 8 in Period 4, 5 in Period 4-5 and 6 in Period 5-6. This reflects a declining trend in the proportion of pigs in the later periods of activity at Heybridge. As with the cattle and caprine remains, the greatest MNI value was derived from the teeth in all periods. The same pattern was observed when the data were analysed by context type (Figure 650) and area (Figure 651). The more robust elements (scapula, humerus, ulna, calcaneum and tibia) are again dominant among the post-cranial material and the small elements almost entirely absent.
The explanation given for the cattle and caprine distributions is again valid here. Recovery and preservation are the major contributing factors to the observed pattern rather than differences in disposal practices, and therefore whole animals were probably brought onto site and the bones not selectively disposed of. Most Roman sites display a range of pig elements with no particular emphasis on any particular disposal pattern. This may reflect the fact that pigs were reared on or in the vicinity of most sites and were hence killed, butchered and eaten 'on the spot'. This is particularly true if many householders kept a few pigs in their backyards as has been hypothesised elsewhere (Dobney et al. c1996).
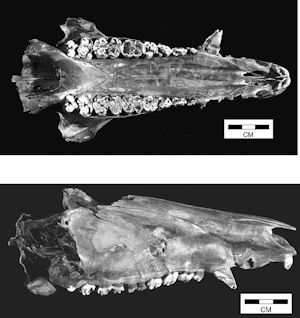
The pig skull recovered from Well Context 20034 (Period 4-5) shows an interesting morphology. The anterior part of the skull is very narrow across the palate and also the nasal profile is long and straight (Figure 652), indicating a rather primitive type of animal.
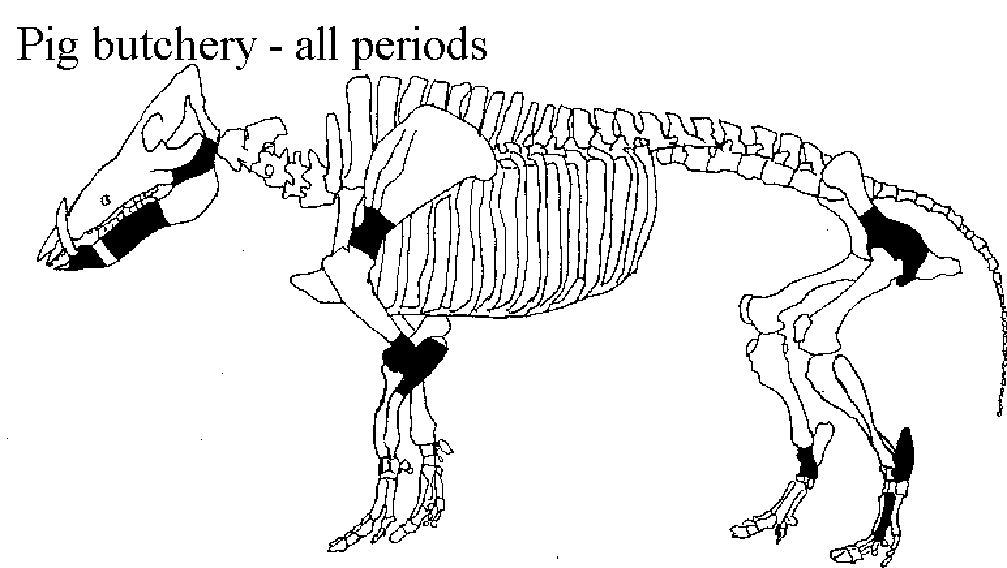
The number of butchered pig bones was very low in all periods. The areas of the skeleton on which butchery was noted are shown in Figure 653 for the whole assemblage. Most of the butchery marks on the post-cranial elements were probably the result of jointing carcasses. The exception to this is a single incidence of a split metapodial, which might have been the result of marrow extraction. The butchery of the mandibles mostly took the form of splitting the two halves of the mandible apart at the anterior end. The most likely explanation of this is to facilitate the removal of the tongue. It is not uncommon for pig bones to display few chopping marks and, as with the sheep carcasses, pigs may have been dismembered using knives rather than cleavers.
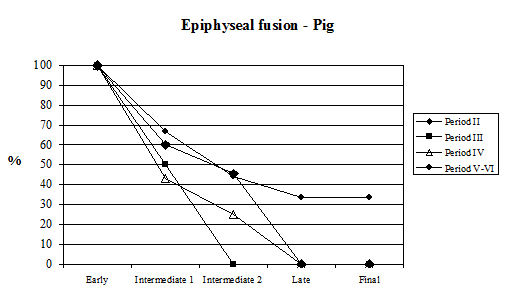
The pig epiphyseal fusion data (Table 139, Figure 654) show a pattern typical for many sites from the Iron Age onwards. Although the numbers in all periods were very small, the striking similarity in the data between periods suggests that the observed trend may be valid. Almost all the bones in the Early category are fused and in all periods except 5-6 all bones are unfused in the Late and Final categories. This suggests that almost all the pigs were killed off between the ages of one year and 2-3 years. This indicates that these were reared for meat and were killed off when their bodyweight reaches the maximum. The presence of a few neonatal and very juvenile bones suggests that breeding of pigs may have taken place on the site or in the immediate vicinity, as there were likely to be new-born casualties. The fact that most of these bones were articulated favours this idea rather than their consumption as suckling pigs.
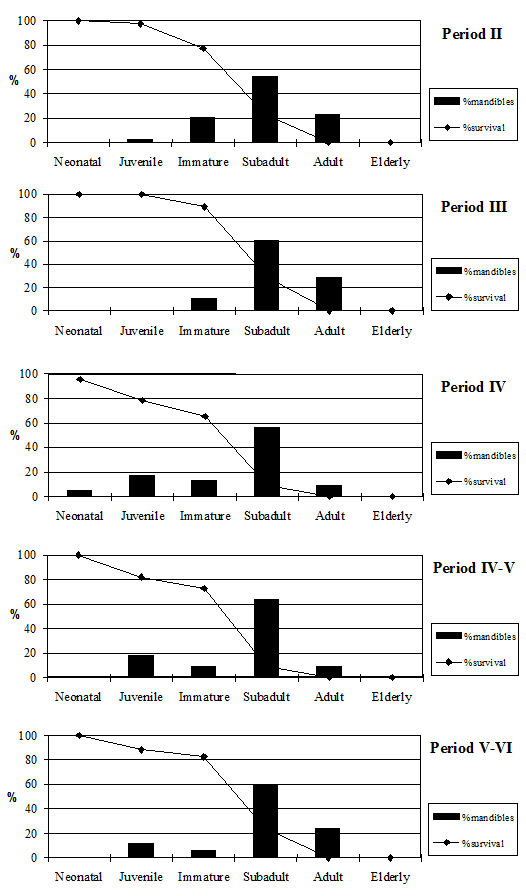
The tooth wear stages were analysed using O'Connor's (1989) categories and the results shown in Table 140 and Table 141 and Figure 655. No mandibles from elderly individuals were recovered and only one from a single neonatal individual was recovered from Period 4. The highest percentage of mandibles in all periods was attributed to the subadult category. However, there appears to be a subtle shift between Periods 3 and 4 towards slightly younger individuals. In general, there are more adult individuals in Periods 2 and 3 and more juvenile individuals in the later periods. This suggests that at around the middle of the 2nd century AD there was a slight shift in the age at which pigs were killed. The presence of younger individuals in the later periods at Heybridge probably goes hand-in-hand with the size increase (see below) suggested for this period. Larger pigs were probably faster growing, and could have been slaughtered earlier.
The practice of killing off pigs before they reach skeletal maturity has been observed at many sites of both Iron Age and Roman date. There is some variation in how young the animals were slaughtered. Most Late Iron Age sites such as Dragonby (Harman 1996), Owlesbury (Maltby 1987), and Skeleton Green (Ashdown and Evans 1981) show that most pigs were killed in the immature and subadult age groups, i.e. between 1 and 2 years old, with some older but few younger. This agrees with picture from Heybridge during this period. For Roman assemblages there is some variability between sites. At Colchester (Luff 1993) there is an increase in juvenile individuals when the city was rebuilt after the Boudiccan revolt of AD 60/1, suggesting a possible increase in the Roman influence on the diet. However, at Roman Exeter the reverse is true, the highest number of juvenile individuals is associated with the initial military garrison.
The General Accident site assemblage from York (O'Connor 1988) contained a number of juvenile individuals suggested to be suckling pig, as well as large numbers of immature and subadult individuals. At Lincoln (Dobney et al. c1996) the early to mid-Roman period follows closely the pattern observed in the later periods at Heybridge, but shows a substantial increase in the number of adult individuals during the 4th century that is not reflected at Heybridge. This suggests that consumption demands varied across the country and also changed through time.

Sexing of the pig remains was undertaken on the canine teeth and their respective alveoli (Table 142, Figure 656). When all the canines were considered together a male:female ratio of 2.5:1 was calculated. However, as the smaller female canines may be biased against by hand-collection the ratio was recalculated using only canines in mandibles and the difference found to be smaller, 1.6:1. When all the canines were considered there was almost no difference between the periods, just a very slight, steady increase in the number of females represented. Looking at just the mandibles the pattern is somewhat different; in Period 3 the numbers are equal and in Period 5-6 nearly equal. The numbers of canines involved is too small to test the significance of these observations but it is suspected they are a product of the small sample size.
Pig assemblages in which young males predominate is the pattern seen on all archaeological sites at which pigs were consumed as the surplus males would be the first to be culled. The presence of some young females can also be explained as the surplus not required for breeding purposes. Looking at the ages of the female pig mandibles, some fully adult individuals were present suggesting that pigs were bred in or near the Heybridge settlement as well as being consumed there. This links with the articulated neonatal skeletons discussed above.
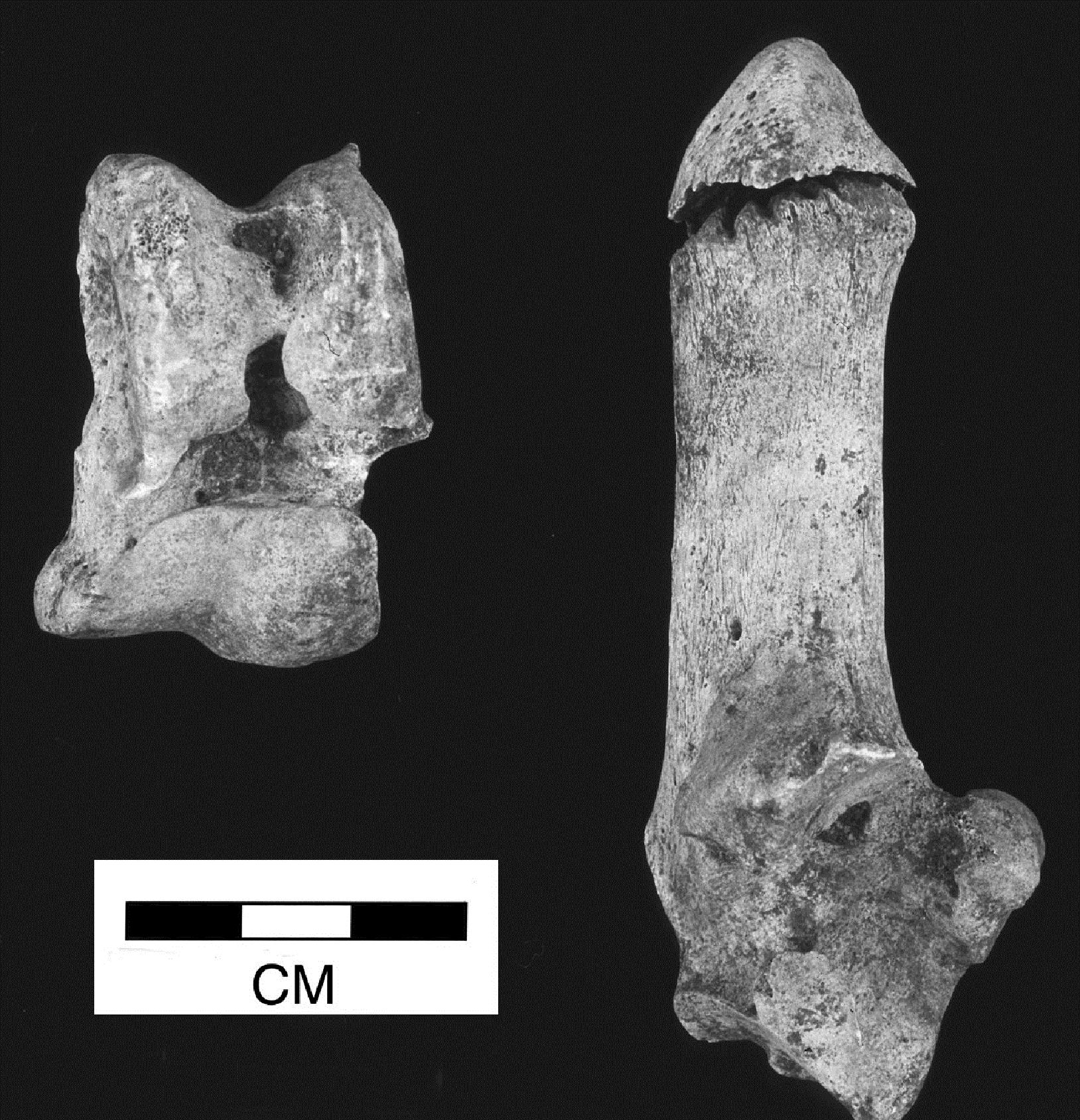
Five pathological pig bones were recorded, two of which are closely related and probably derive from the same individual. A calcaneum (context 13094, Group 432, Period 4) exhibited a possible healed fracture of the anterior, distal part, splaying of the distal articular surface and new bone formation above the sustentaculum (Figure 657). An astragalus in the same context displayed splaying of the anterior articular surfaces and also slight eburnation. If these are from the same individual, the fracture to the calcaneum may have subsequently caused the changes to the astragalus in compensation. The other three pathological specimens were metapodials. The first of these, a metacarpal (context 4899, Group 749, Period 3) appeared to have a slightly abnormal distal end. The unfused distal end appeared splayed with extra bone growth around the margins. Also the whole bone appeared short and flattened. The second metacarpal (context 5214, Group 442, Period 5-6) showed a well-healed and remodelled fracture of the lower part of the shaft, possibly a greenstick fracture. The distal end of the metaphysis and the epiphysis of the metatarsal (context 15233, Group 471, Period 6) were slightly abnormal, possibly as a result of trauma when juvenile or a slight misalignment while fusing.
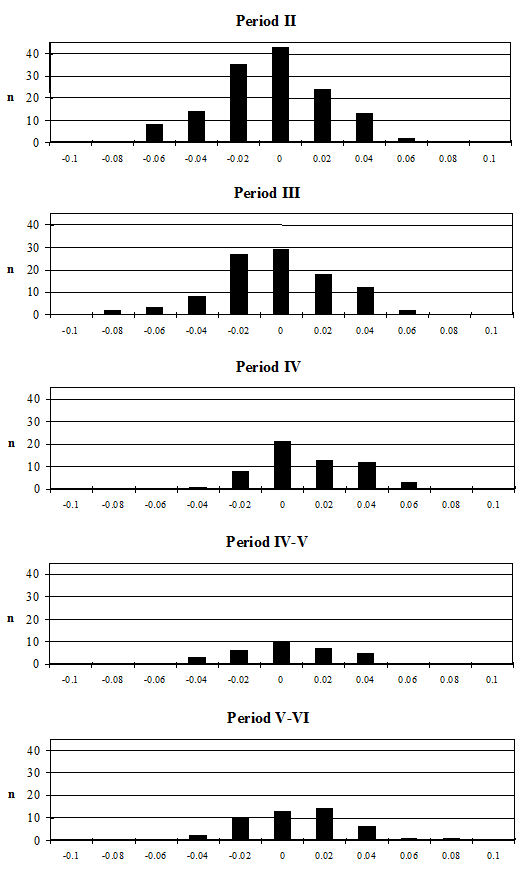
As with most archaeological sites, there were rather few measurements of pig post-cranial material, although a reasonable number of tooth measurements could be taken. Table 143 summarises the measurements for which greater than ten cases were recorded (all individual measurements are given in the archive). Looking firstly at the teeth measurements, the log ratio technique was employed using a group of Neolithic pigs as the standard (see methods section). There are no obvious differences between Periods 2 and 3 (Figure 658) and the size range appears symmetrical around the standard value (0), suggesting that the teeth of the Heybridge pigs do not substantially differ in size from those of Neolithic pigs. Although there is only a slight visual increase from Period 3 to 4, it is highly significant statistically (Table 144), and it is therefore likely that, like sheep, pigs would have been improved by Period 4. The last two periods present a somewhat more confusing picture, Period 4-5 appears statistically more similar to Period 3 than either of its neighbouring periods, whilst Period 5-6 is statistically similar to Period 4 again. Since there is no clear trend over time, we have to be cautious about the interpretation of these data particularly because the statistic, though useful, was not applied in an orthodox way.
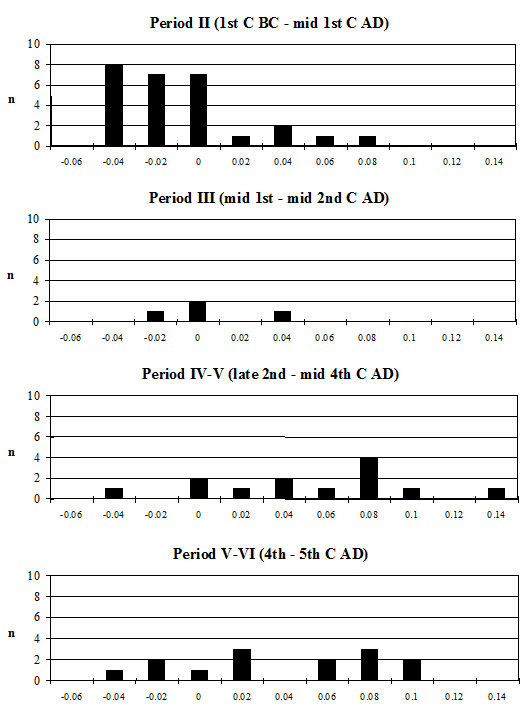
As there were so few post-cranial measurements, all were combined using the log ratio technique to gain an adequate number of cases for analysis of at least some periods (Fig 659). Period 2 shows that most of the post-cranial material was slightly smaller than the standard. The differences between Period 2 and Periods 4-5 are marked, even visually, and as would be expected are highly significant statistically, which also provides more credibility to the trends observed in the teeth. Period 5-6 is similar to Period 4. This suggests that some time after Period 2 the size of the pigs at Heybridge clearly increased. Unfortunately, there were few measurements in Period 3, but these do seem to be small also. Therefore, an increase in the 2nd century AD seems to be likely.
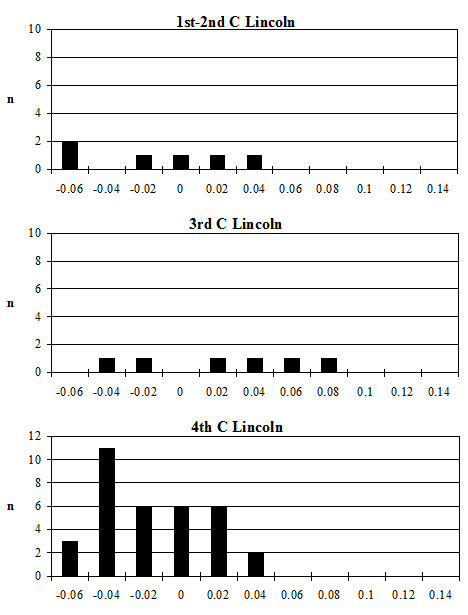
The only comparative data for Roman pigs came from Lincoln (Dobney et al. c1996) as most sites produce too few measurements. Only the 4th century provided a large sample of material, which is comparable in date with Period 5-6 at Heybridge (Figure 660). The differences are very pronounced, with the Lincoln material being small in comparison to both the standard and the Heybridge material. The difference between the Heybridge and Lincoln material is highly statistically significant and in fact the 4th-century Lincoln pig bones are more similar to those from Heybridge Period 2, that to the contemporaneous material. Few measurements are available for 3rd-century Lincoln, but these seem to be larger and similar to those from contemporaneous Heybridge. A size reduction therefore seems to occur between the 3rd and 4th century in Lincoln. This replicates a pattern already seen for cattle.

Instances of wild boar on Roman sites are very rare (Luff 1982) and most identifications are based on the biometrical evidence (Maltby 1979). This is also the case at Heybridge, where three possible wild boar fragments were identified, all from Period 4 deposits. The first was a calcaneum (context 4427, Group 739) with a greatest length of 107.9mm. This shows up as the large outlier in Period 4-5 of Figure 660. The second was a scapula (context 6672, Group 379) with a GLP measurement of 46.2 and SLC of 32.2mm (Figure 661). The last was a 1st phalanx (context 6121, Group 556, Period 4) with a greatest length of 51.4mm. The measurements are all considerably larger than the (admittedly small) wild boar in the BZL reference collection and are also similar to, or larger than, the Turkish boars used in Payne and Bull's study (1988).
A total of 169 equid fragments were recovered from Heybridge. For the Roman period this represents 3% of the total of cattle and equid bones. In the Iron Age the percentage is only slightly higher (5%). A survey of 190 Roman sites in central England reveals that, on average, equids represent 5% of the total of equid and cattle bones. The ratio between the two taxa is higher on rural sites (1:10) and lower in towns (1:25), whereas military sites produce an intermediate value (1:20). These differences are more likely to be due to waste disposal patterns than to economic or social factors. It has to be remembered that the zooarchaeological evidence does not provide information on animals that were kept on site but died and/or were disposed of elsewhere. This could have been the case for most non-food animals in urban areas. It is interesting that at Elms Farm, in all periods, the frequency of equid bones is much more similar to what is found on urban than on rural sites.
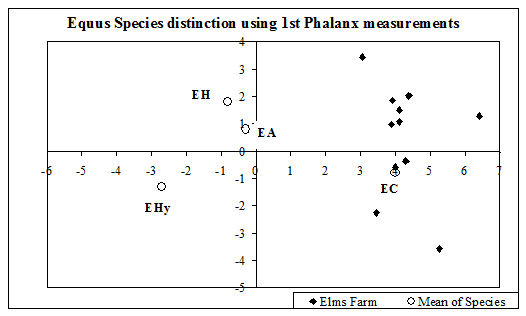
A wide range of elements were present (Table 145) but the most numerous were loose teeth, 1st phalanges, metapodials, tibiae and astragali. Many of the loose teeth were incisors and were therefore of no use in determining species. Even for the remaining loose teeth only a few were precisely identified and only single mandible and maxilla fragments were present in the whole assemblage. As there were so few teeth that could help determine the species of the Heybridge equids it was decided to try a different approach using the unpublished work of Simon Davis on separating Equid first phalanges using multivariate analysis. Twenty-two first phalanges were recovered, but for only eleven of these could all six required measurements be taken. A plot of the Heybridge measurements on the same axes as in Davis (Figure 662), shows that most of those from Heybridge fall within the range of E. caballus (horse). However, a few lie outside the range of the horses used in the study but seem to be nearer this group than the other equid groups. As all the E. caballus specimens used in Davis's study were small Przewalski ponies, there is the possibility that more 'refined' and/or larger types of horse and pony may show differences in conformation. The likelihood that the equid remains at Heybridge are not horses is slim, so for the rest of this report they are all assumed to be horses.
Most of the horse bones were disarticulated finds but one articulated part skeleton was recovered from Well 6641 (context 6640, Group 579, Period 5). The fifty bones belonging to the same individual were mostly limb elements, but a few teeth and axial bones were also present. The individual represented was an adult (all the epiphyses were fused) and stood about 14 hands (average for individual bones was 1402mm) at the withers. The only slight pathological condition noted was that two of the first phalanges had slightly exaggerated muscle insertions on the shaft area, possibly the result of work-related stress.
Almost all the bones and teeth represent adult individuals as the epiphyses were fused and the permanent teeth were in wear. The exceptions were an unfused calcaneum from Period 4, and unfused distal tibia from a Period 5 deposit and a fusing distal tibia from Period 6. In addition a slightly porous terminal phalanx was recovered from a Period 4-5 deposit. All these indicate animals below the age of three years. The permanent incisors were all worn but mostly not excessively so. One upper 3rd incisor showed the characteristic 'hook' at the posterior edge of the biting surface that appears when a horse is in its seventh year (Goody 1983). Another incisor had very little of the infundibulum left, which probably means the animal was at least 15 years old. The absence of very young individuals and only a few immature bones suggests that horse breeding was not taking place on site, and that most of the horses present were working animals in their prime and onwards.
The presence of butchery marks on horse bones is always a slightly controversial subject in view of the modern (British!) distaste for eating horsemeat and has been discussed at length on many occasions (e.g. Lauwerier and Robeerst 1998). In most 'Romanised' parts of the Empire, horsemeat was only eaten in emergencies (Tacitus Annals II, 24 and Histories IV, 60; quoted in Luff 1982). Ten horse bones from Heybridge showed evidence of butchery (all except one from post-conquest deposits), which equates to 6% of the equine bones recovered. This figure is higher than that observed on sites in the Romanised Netherlands (Lauwerier 1988) but is still substantially lower than that for the cattle. Given the Roman aversion to horsemeat and the low percentage of butchered bones, in contrast to cattle, it is likely that horses were rarely, if ever, used for human consumption at Heybridge.
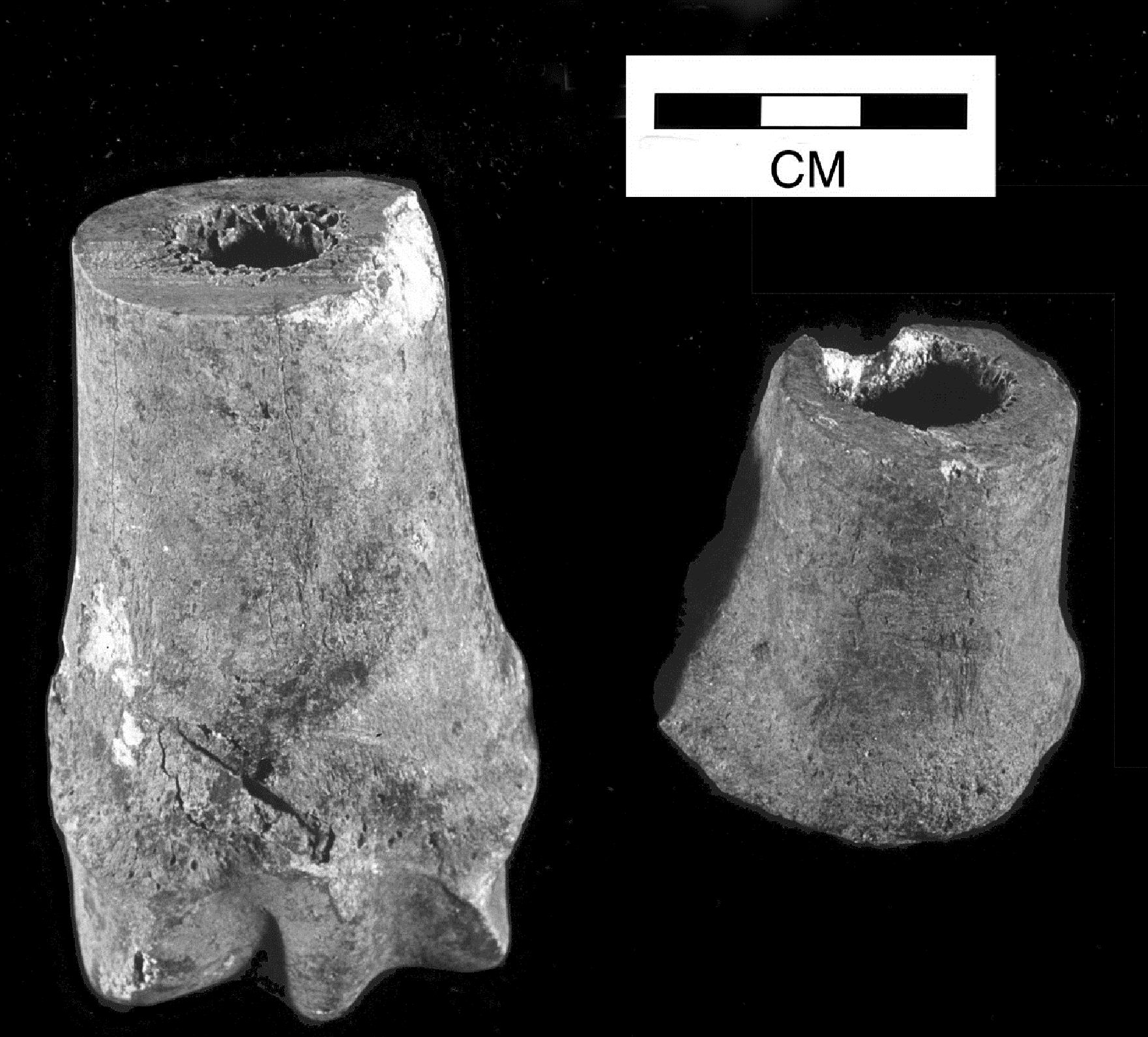
Of the bones bearing cut marks, four were metapodials and phalanges with horizontal cuts, probably indicating that the animal was skinned. A radius and tibia had been split longitudinally, possibly the result of marrow extraction. Other bones included two scapulae, which had been chopped longitudinally, and an ulna that had horizontal chop marks just above the articular surface. These marks might be the result of butchery for meat, as the locations tend to rule out other factors, but possibly for consumption by dogs rather than humans. This was the hypothesis put forward by Lauwerier (1988) for the presence of butchery marks on horse bones from various Roman sites in the Netherlands and it also seems to fit the evidence here. In addition, two metapodials were recovered that had been sawn horizontally through the shaft (Figure 663). This may indicate that these fragments were the waste product of bone working. This type of bone-working waste is more commonly found in medieval deposits, though sawn bones have occasionally been found in Roman deposits too.
Ten pathological horse bones were noted, mostly associated with joint problems. These included a metacarpal (context 14784, Period 2) with splints - the accessory metacarpals fused to the main bone with accompanying extra bone growth. A second metacarpal (context 14558 Period 6) displayed extra bone growth around the accessory metacarpal joints, possibly the beginnings of splints. Splints can be caused by trauma to the area, by stress to the joints or old age. Spavin is another pathology of this area consisting of the fusing of the tarsal (or carpal) bones and with a similar aetiology to splints. The articular surfaces are unaffected but in severe cases the joint in completely bridged by bone growth, rendering it immobile. A single case was noted from Heybridge; two completely fused tarsals from context 6640 (Group 579, Period 5). Lower down the limbs, three first phalanges (contexts 4924, Group 739, Period 4 and two in 6640, Group 579, Period 5) were affected by areas of bone growth, taking the form of very exaggerated muscle insertions on the shafts. A single third phalanx (context 14613, Group 722, Period 2) was also affected in this way with bony growth extending from the articular surface and around the tendons. All these are most likely to be stress (perhaps due to the use of the animals for traction) or age related.
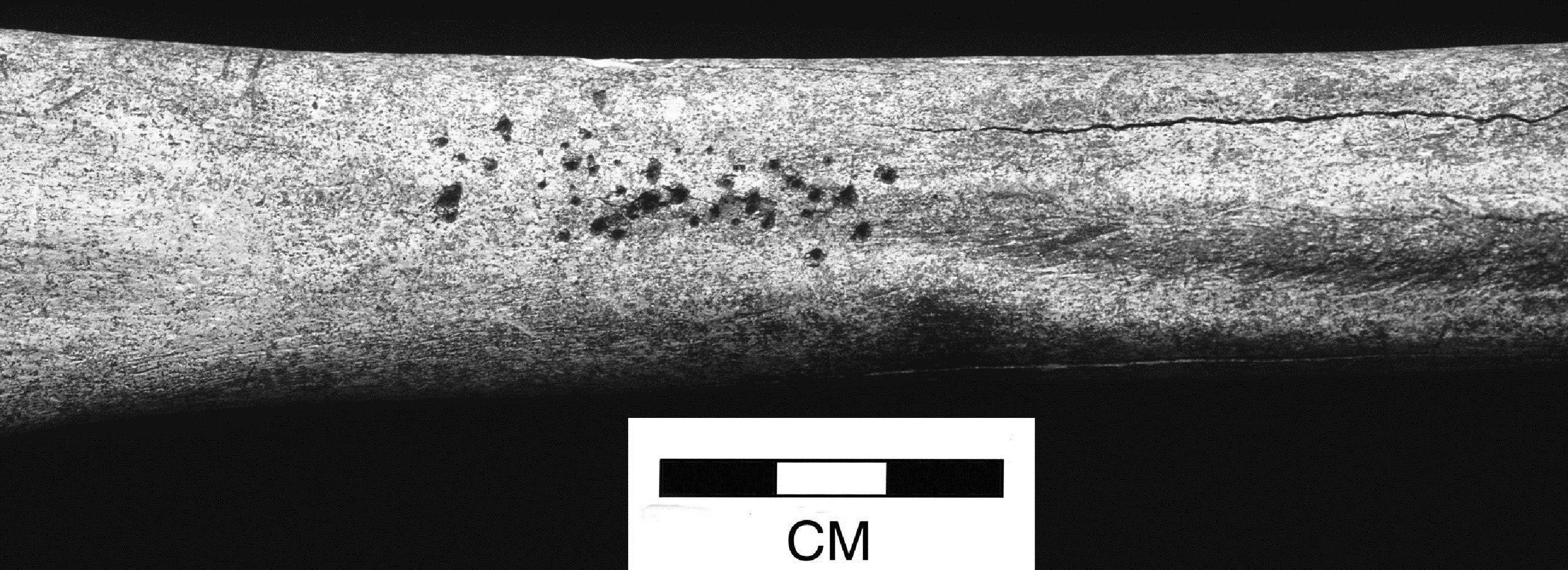
A single tibia (context 9415, Group 2085, Period 2) displayed a trauma to the bone when immature. A line was visible across the distal articulation, possibly indicating a trauma to the epiphysis prior to fusion resulting in the epiphysis not fusing properly. A lower cheektooth (context 4993, Group 750, Period 2) showed evidence of enamel hypoplasia with both linear and isolated defects about halfway down the crown of the tooth. In addition, a metatarsal (context 5228, Group 457, Period 5-6) was noted that had a patch of small round holes in the centre of the posterior shaft. Their appearance is similar to the holes the point of a pair of compasses makes when stabbed into wood. Two explanations offered include post-mortem parasite attack or the use of the bone as a rest while undertaking some activity involving making small holes (Figure 664).
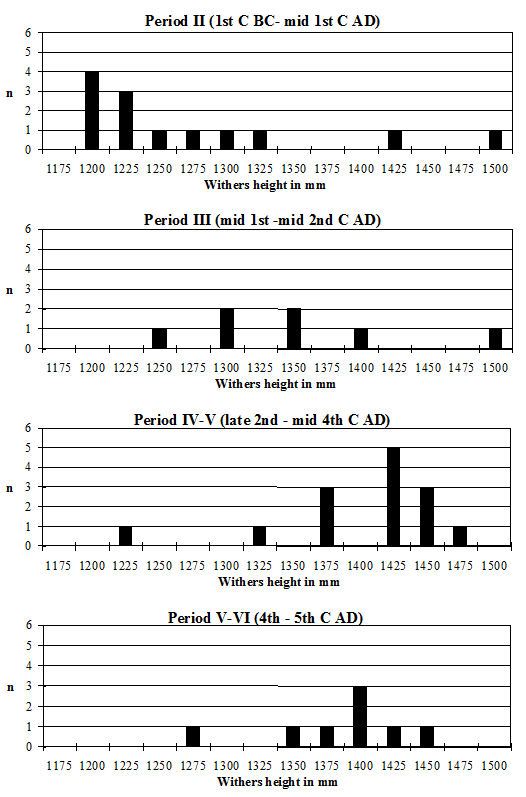
The post-cranial biometrical data for horses was more numerous than for the pigs but no elements produced more than ten measurements within any one period or indeed overall, so the individual measurements are given in the archive. As with the pigs and sheep the paucity of measurements meant that techniques enabling data to be combined had to be employed. For horses in particular one standard way of doing this is to use estimated withers heights. For the Elms Farm material, the factors of Vitt (1952) were used for the main long bones and that of Johnstone (1996) for the first phalanx. Table 147 gives the estimated withers heights by bone. Figure 665 shows the data by period and it is very easy to see that the Period 2 horses were considerably smaller than later ones (apart from two large individuals). There is also a hint that the horses got smaller again in Period 6. The mean height of the Iron Age horses was 1260mm (12.2 hands) and the Roman horses was 1376mm (13.2 hands) giving an increase of just over 10cm or one hand. Looking at the modal classes the size increase is more obvious, from under 1200mm in the Iron Age to just over 1400mm in the mid-Roman period. In real terms this is a considerable increase in the size of the horses, from an animal of similar height to a modern Welsh Mountain pony to that of a large New Forest pony.
Table 146 gives a summary of the withers height data from Heybridge and other sites. It can be seen that the Period 2 data fits towards the upper end of the range from other Iron Age British sites and are somewhat smaller than those seen on the continent at around the same time. For the Roman period the Heybridge mean is very close to that for the collection of British sites and fits well with the data from sites in the locality. The slight oddity is that the horses found at Colchester were generally smaller than their counterparts elsewhere in Essex. As in the Iron Age, the continental material is somewhat larger, but these remains were from high-status and military settlements, which may influence the size of the horses found there (Lauwerier and Robeerst 1998).
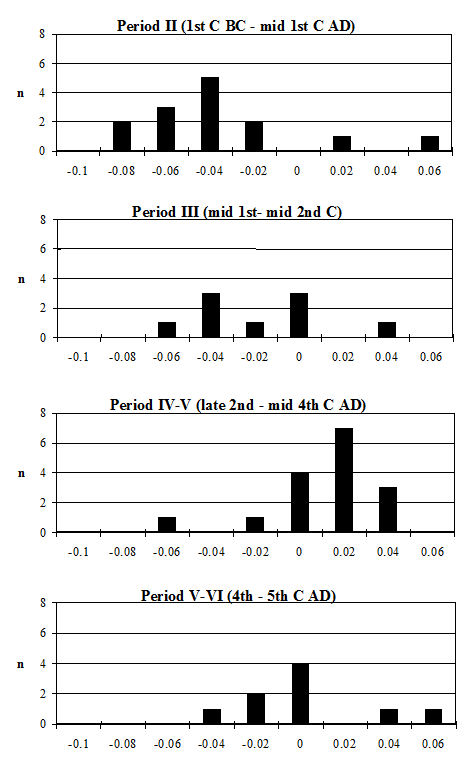
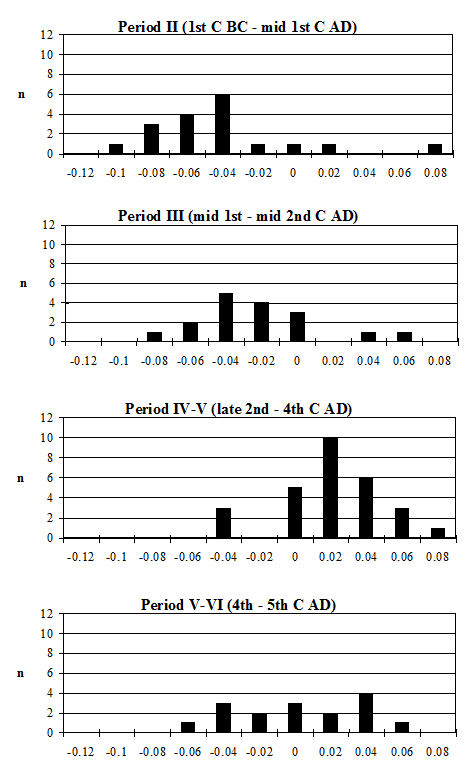
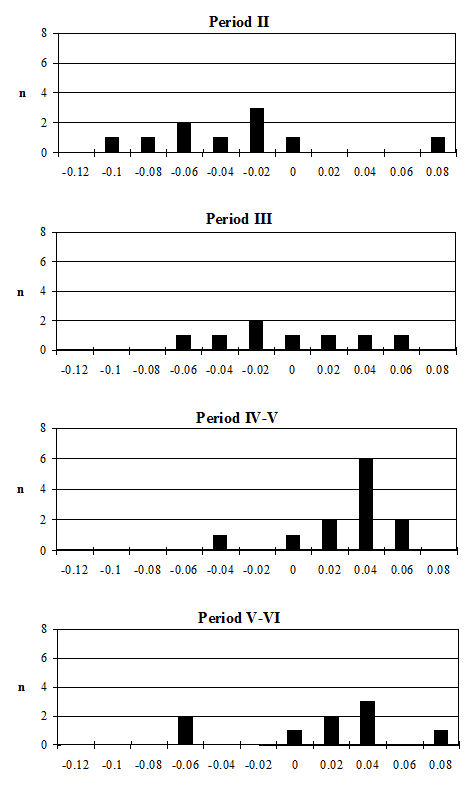
As with the other species the log ratios of the length, width and depth measurements were calculated as another means of combining the data. The standard used for comparison was the means calculated from measurements collected from a series of Roman sites in Britain (Johnstone 1996). The size increase seen in the withers height reconstruction data is also evident in the log ratio data (Figures 666, 667 and 668). As there are more cases in all periods using this method, it is possible to see that the main size increase occurs between Periods 3 and 4 as has been observed for all species except the cattle, for which it occurs earlier. In all three dimensions there is no significant difference in size between Periods 2 and 3 and also no difference between Periods 4-5 and 5-6 (t-tests in Table 148). However, there is a significant difference between Period 3 and 4-5 in the length and depth measurements and a highly significant difference in the width measurements. The withers heights (Figure 665) and log ratios of lengths (Figure 666) and widths (Figure 667) show a visual decrease in size in Period 5-6. Although this change is not borne out by the statistical tests (possibly the result of small sample sizes) the size decrease in the very late Roman and Early Saxon period is consistent with trends across Britain (Johnstone 1996).
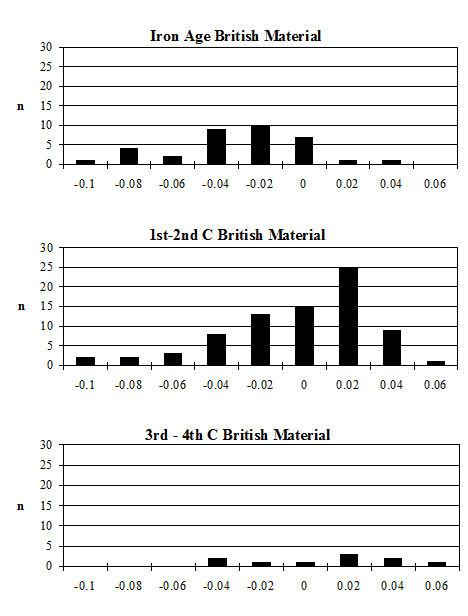
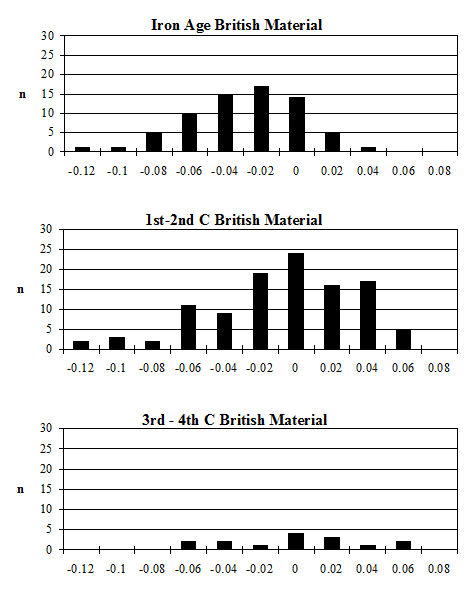
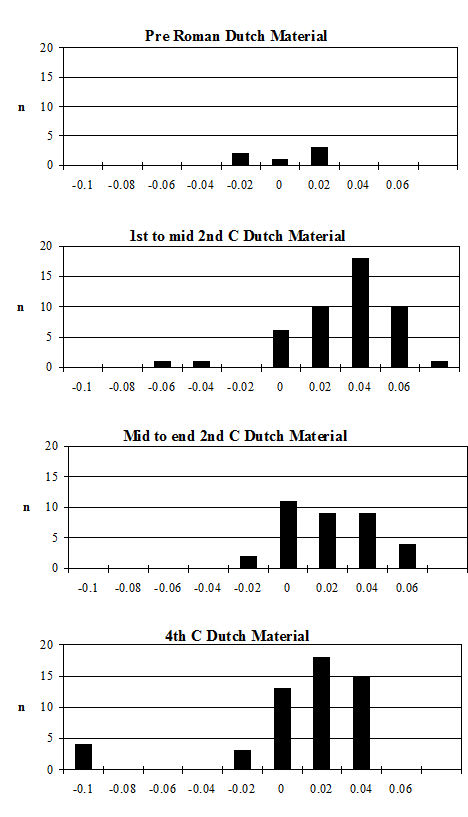
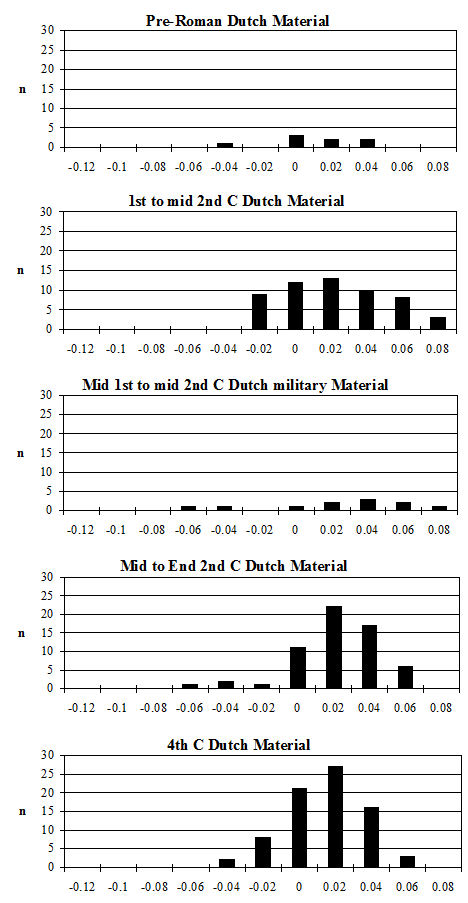
Looking at the log ratio data in a wider context (Figures 669, 670, 671 and 672) the same patterns emerge as for the withers height data. The Heybridge Period 2 material fits well within the British Iron Age data for both length and width measurements, and is again significantly smaller than pre-Roman Dutch material. The Period 3 material fits within the range for the 1st and 2nd-century British material with no significant difference and is (highly significantly) smaller than Dutch material for the same period. For the later periods at Heybridge the size of the horses does not significantly differ from either the combined British data or the 4th-century Dutch data. This suggests that while British horses were smaller initially, the Roman influence on horse breeding increased the size of the individuals to match their continental counterparts by the middle to Late Roman period.
Of the 240 canid remains, most were assumed, on the basis of their size and morphology, to be those of the domestic dog. A few were of similar size to fox bones and thus could not be identified with certainty. Most of the dog bones (197) were from articulated whole and part skeletons (Table 120). Several of the more complete skeletons were recovered from one feature, Well 14984 (Group 710, Period 4-5, Area L). Two contexts from this well (20034 and 14939) contained articulated dog remains in addition to cattle and pig skeletons. Although the remains had originally been articulated, the connection had not been kept at the excavation stage, which meant that most elements could not be attributed to individuals. The major limb bones were separated out on the basis of size and a minimum of four individuals were recognised in context 20034 (Group 713) and two in context 14939 (Group 713). All the bones had fused epiphyses and all the mandibles had permanent teeth, suggesting that all the individuals were adult at the time of death.
The other almost complete individual came from context 6152 (Group 552), a pit fill (Area H, Period 4). Parts of this were recovered by hand-collection and the remainder by sieving. This individual - an adult male - indicates what could have been retrieved from other individuals where sieving did not take place. Several bones of this specimen were fractured and other pathologies were also present. These comprised a tibia with healed but misaligned mid-shaft fracture, with large callus formation around the fracture site. The ulna was also fractured but remained unhealed and was enlarged with new bone growth that fused it to the radius. The new bone growth may also be the result of infection. The distal end of the same radius displayed exostoses around the joint, probably a result of the fractured ulna. Also one jaw displayed a dental anomaly, either pathological or congenital in origin. The lower first molar was impacted into the jaw and the surrounding area of bone very swollen in appearance. The tooth is hardly worn but must have been fully erupted as the second molar is in wear. Possibly a large abscess had formed below the tooth and it dropped back into the jaw. This individual seems to have had a rough life and some of the fractures that we noted, being unhealed, may have led to his death, owing to accident or mistreatment. A similar example of a burial of an apparently mistreated animal was found at Silchester Insula IX (Atkinson, pers. comm.).
Other pathologies were occasionally found on loose bones and are recorded in the archive.
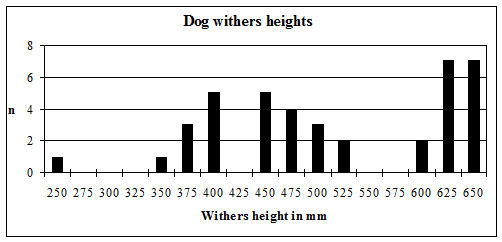
Measurements were taken on the long bones and also on the mandibular and maxillary teeth. The tooth measurements all showed that they were from domestic dogs, as the proportions of the teeth to the length of tooth row showed quite a crowded jaw and the upper 1st and 2nd molars showed a dog-like ratio (Clutton-Brock 1969). The lengths of the long bones (all from Roman deposits) were converted to shoulder height estimates using Harcourt's factors. These gave a range of 249 to 637mm (Figure 673). This range is similar to the range of sizes seen at Colchester (Luff 1993) and is also within the range for Roman dogs given by Harcourt (1974). The smallest withers height (249mm) was obtained from a tibia with a distinct curvature. This has been observed in many small dog bones from Roman sites and is characteristic of what has been termed the small lap-dog type. Most of the dogs were between 350 and 525mm at the shoulder but a few reached over 600mm in height. The size and shape of these larger bones (from context 20034) were similar but slightly more robust than the greyhound in the BZL reference collection, suggesting they may represent hunting-type dogs.
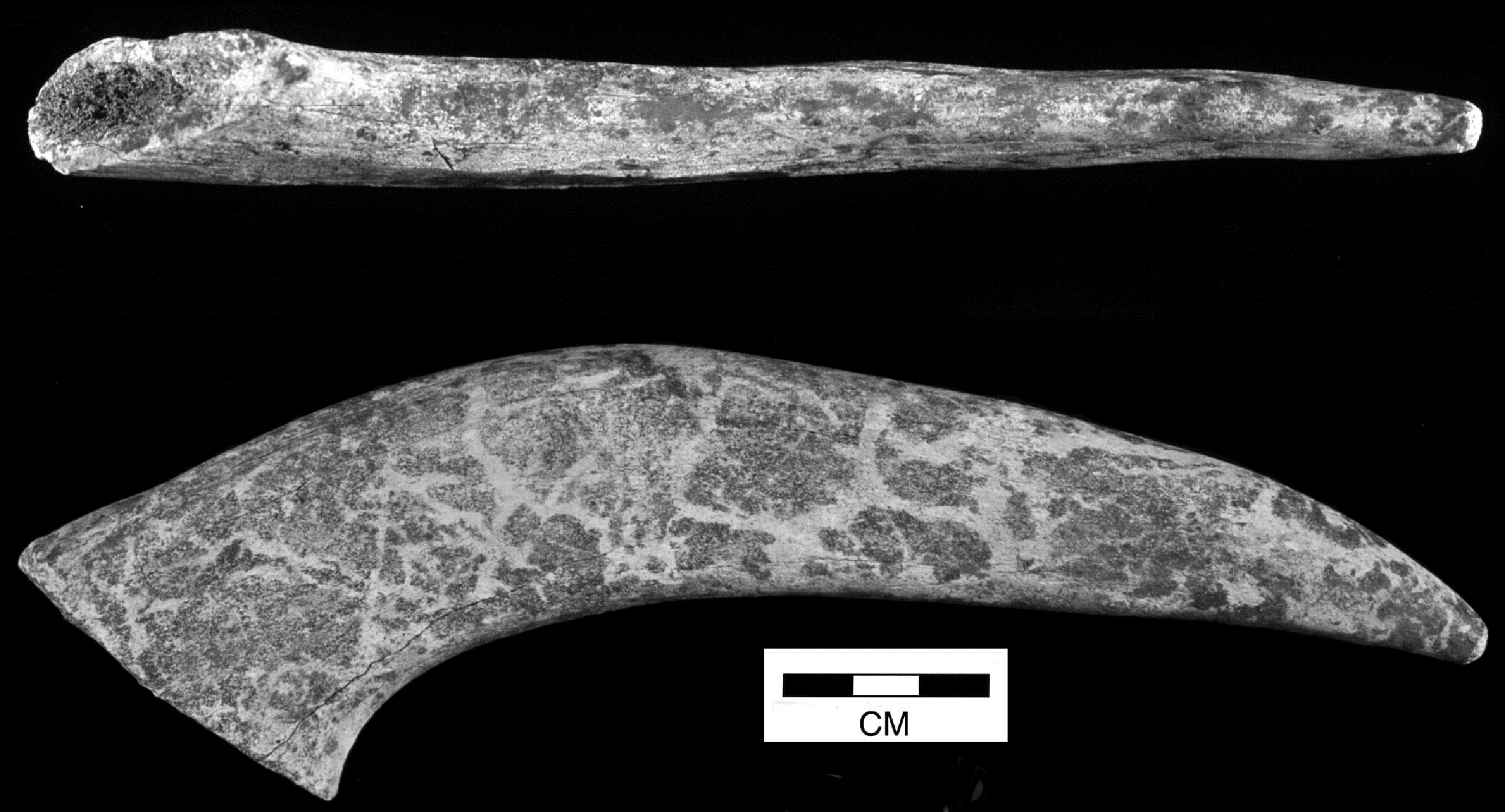
Of the larger wild mammals deer provided the greatest number of fragments with sixteen red deer (Cervus elaphus L.) and a single roe deer (Capreolus capreolus (L.)) in the countable section. The single roe deer fragment was a radius from a Period 3 deposit. The red deer remains were mostly from Period 4 (nine fragments). Most bones belonged to non-meat bearing elements i.e. butchery waste; teeth, mandible, axis, distal radius, distal tibia, calcaneum, metapodials and phalanx, the exceptions being a proximal ulna, distal scapula and distal humerus. The mandible was from an old individual as the teeth were very worn. A single pathological deer bone was recorded; a first phalanx (context 4707, Group 3034, Period 4) with extensive splaying and very slight eburnation of the proximal articular surface. This find should remind us to be cautious when interpreting such pathologies - when found on domestic animals - as a consequence of traction stress. A single antler fragment (context 14558, Group 722, Period 6) showed a very flattened profile (Figure 674), which may be natural variation as there is no evidence of any pathological condition.


In addition, a substantial number of antler fragments were present in the non-countable section. Three were identified as roe deer antlers, forty-one as red deer, two as probably red deer and ten as red/fallow deer. However, the last group are more likely to be red deer, as the existence of fallow deer in Britain in the Roman period is still under debate and, if present, were very scarce (Sykes 2010). More than half of the antler fragments (thirty-four out of fifty-six) were located in Feature 14529 (Group 722), a well dated to Period 6 (latest Roman-Early Saxon). Almost all of these fragments were antler-working waste i.e. the pieces of antler less favoured for working, such as the burrs, junctions and tine tips (Figures 675 and 676). While antler products are known from the Roman period, they become far more prevalent from the beginning of the Saxon period onwards (MacGregor 1985). This hints at the possibility that the deposits containing this antler waste are more likely to date to the Early Saxon period rather than the Late Roman, and hence could be associated with the nearby Sunken Featured Building.
Single bones of hare (Lepus europaeus Pallas) and badger (Meles meles (L.)) were recovered. The hare bone, a scapula (context 11139, Group 676, Period 5), may well represent a hunted food resource, but a single bone is insufficient upon which to base any assumption. The badger element, an ulna (context 4705, Group 4016, Period 4), is unlikely to represent a food resource and is more likely to indicate that scavengers were active within the settlement or the hunting of this animal for its fur.
Forty-five recordable small mammal bones were recovered from Elms Farm, all except one from the sieved material. The species represented included small voles, mice, shrews and water vole. The mouse species present are commensal and are commonly found in close association with human habitation. The small vole species tend to inhabit grassland environments suggesting that at least part of the area immediately adjacent to the settlement was relatively open. Although the water vole fragments were recovered from central areas of the site, the presence of a watercourse on the north side of the site suggests these were natural/accidental deaths.
A total of eighty-eight chicken bones were recovered from Elms Farm, seventy-four by hand collection and the remainder from sieved samples. The distribution of chicken bones across the site was very unevenly spread with sixty-two of the bones being recovered from Area J (the temple precinct). Fifty of these were from Period 4 deposits, representing a minimum of seven individuals. Looking at the distribution of these bones, almost all were recovered from two pits (Group 432: Feature 5394 - Fills 5392 and 5393 and Feature 13084 - Fills 13071, 13072, 13083 and 13094) near the south-west corner of the temple precinct. The proximity of this relatively large group of bones to the temple complex suggests that they may well have been sacrificial offerings, as hypothesised for the sheep mandibles above. Only two bones were recovered from Period 2 deposits. This is not all that surprising as chickens are rare in Britain before the Roman period. The contexts that produced chicken bones in Period 2 (4506 (Group 278) and 17282 (Group 330) may well be post-conquest.
All the larger and more robust elements (coracoid, humerus, ulna, femur, tibiotarsus and tarsometatarsus) were represented evenly across the site, spatially and temporally; the lack of scapulae and carpometacarpi may have been caused by either preservation or recovery bias. A few pathological bones seem to have little potential for archaeological interpretation and are described in the archive.
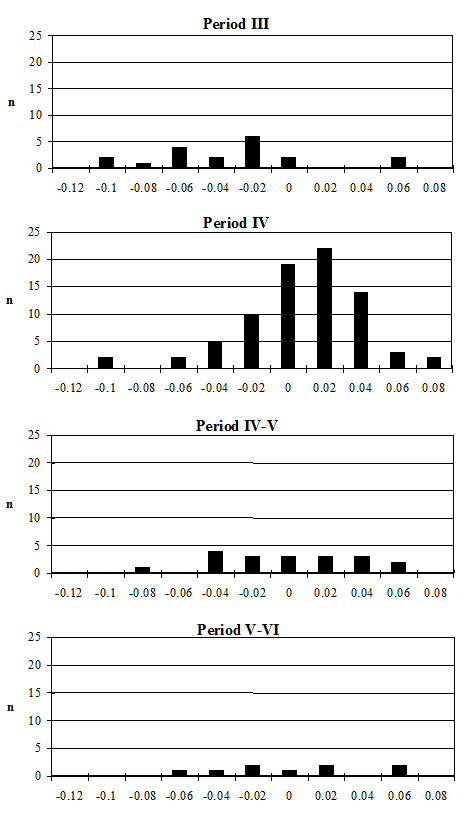
| Measurement | Site/s | Comparison between: | T value | Probability | Significance |
|---|---|---|---|---|---|
| All Log ratios | EF | Period 3 and 4 | -4.333 | 0 | ** |
| EF | Period 4 and 4-5 | 0.816 | 0.4167 | N | |
| EF | Period 4-5 and 5-6 | 0.152 | 0.8806 | N |
As with most of the mammal species, the number of measurable bones was rather limited so the log ratio technique was employed to make the most of the available data. The size increase between Periods 3 and 4, which has been seen in most of the other animals, is also apparent in chickens (Figure 677) and is statistically highly significant (Table 149), while there is no significant changes between the later periods. In terms of absolute size the Period 3 birds were slightly smaller than the standard (a female Old English Gamebird from the Environmental Archaeology Unit York reference collection), whereas the later material was slightly larger. In relation to other sites, the Heybridge chickens were about the same size (or slightly larger) than the corresponding periods at Lincoln (Dobney et al. c1996). The range of tarsometatarsal greatest lengths given for the Colchester Roman material (Luff 1993) encompasses that from Heybridge and also the even narrower range from Great Holts Farm (Albarella 1997). There were too few measurements to address the question of sex of these individuals biometrically, the only guide being the presence or absence of spurs. Of the twelve tarsometatarsi present, six had spurs present, one had a spur scar (B. West 1985) and on one there was definitely no spur. Of the remaining four, two were juvenile individuals and the other two were unclear (area missing). This suggests that seven were likely to have been male and one probably female.
A few bones of geese and ducks represent the only other (potentially) domestic birds found at Elms Farm. These are distributed across all periods, with the exception of Period 2. Although a few post-conquest contexts may belong to Period 2, this period is mainly dated to the pre-Roman Iron Age. Therefore, the absence of geese and ducks is unsurprising as these species may have not been introduced to the country in their domesticated form until the arrival of the Romans. The total number of bones is too small to try to determine any spatial or chronological trend in the distribution of these two taxa.
The domestic or wild status of these birds is very difficult to determine as there are no reliable morphological characters that allow us to distinguish the bones of domestic geese and ducks from those of their ancestral species, the greylag goose (Anser anser) and the mallard (Anas platyrhynchos). Distinguishing them from other, closely related, wild species may also be problematic. Nevertheless, bones of domestic forms tend to be larger. All goose bones from Elms Farm are comparable in size to greylags and - as far as we can tell - might therefore be domestic. Though a fair amount of duck bones are similar to those of mallards, others are smaller and more similar in size to a wigeon (Anas penelope) (or other wild duck in the same size range).
Domestic geese are known to have been kept by the Egyptians, Greeks and Romans (Clayton 1984a), but the status of the duck is more uncertain. The species may have been domesticated in the Far East as early as the 3rd millennium BC, but the Romans may have simply fattened captive birds (Clayton 1984b).
Of 190 Roman 'period sites' (i.e. different periods of different sites) surveyed for central England, only about sixty have produced bones of either geese or ducks. Some bones have been identified as definitely 'wild goose' in only two of these 'period sites'. Wild ducks were found at four sites, though these do not include very small ducks, such as teal and garganey, which are easy to separate from the other species, and which in this report too are discussed separately. Absence of anatid bones can result from the small size of some assemblages, to poor recovery or to a lack of a tradition in keeping or catching these birds. Only a handful of sites includes material recovered through sieving. Among hand-collected assemblages that include either taxa, twenty have produced only goose bones, thirteen only duck bones and twenty-four bones of both species. Of the latter, duck is predominant in fourteen cases, goose in three; four assemblages have equal numbers of the two taxa and for three assemblages bone counts are not available.
Sites located in Essex seem to follow a similar trend. Three have only goose bones and two only duck bones. These latter include the Late Roman temple at Caesoromagnus that has produced a single duck bone interpreted as wild (Luff 1992). The only site with bones of both taxa is Colchester, where duck is almost twice as common as goose (Luff 1993).
The apparent oddity in the above statistics is that goose seems to be more commonly found across different sites, but when both taxa are present duck is almost invariably predominant. We are inclined to believe that this is due to a recovery bias, as in some sites where the rate of collection may have been particularly poor, it is more likely the smaller duck bones were overlooked. This is confirmed by the fact that in the only four sites to produce bird bones from sieved samples, duck is invariably more common. The sites in question are Gorhambury, St Albans (Locker 1990), Great Holts Farm, Essex (Albarella 1997), Scole-Dickleburgh, Norfolk (Baker 1998) and Bonners Lane, Leicester (Baxter 2004). A more general survey of all Roman Britain (Parker 1988) indicates a prevalence of duck over goose records.
All in all, the evidence indicates that at Heybridge, as in the rest of Britain, ducks were kept (or caught) in greater numbers than geese. This is in sharp contrast with what we know for the Saxon and medieval period, when geese are by far predominant and duck meat was, at least in some areas, regarded as of poor quality or even unhealthy (Grand and Delatouche 1950). The Roman preference for duck meat ended soon after the Roman retreat from Britain, as proven by the clear predominance of geese in Early to mid-Saxon sites such as Pennyland, Milton Keynes (Ashdown 1993), St Albans Abbey (Crabtree 1983) and, most remarkably, Ipswich (Jones and Serjeantson 1983), West Stow (Crabtree 1989) and Flixborough (Dobney et al. 2007). These last three sites have produced large assemblages of bird bones where duck accounts for less than 10% of the total duck and goose.
The few Late Iron Age sites that have available evidence of ducks and geese (probably wild) follow the same trend as the Roman sites, with ducks generally predominant. These include, among a few others, the Belgic settlement at Bierton, Aylesbury, Buckinghamshire (Jones 1986), and the enclosure at Wardy Hill, Cambridgeshire (Davis 2003).
It will be interesting to try to determine in the future whether this trend is due to the fact that most anatids from Roman sites are wild, in continuation with the Iron Age tradition, or to a Romano-British preference for duck keeping, in contrast with later Saxon and medieval practices. The evidence from Elms Farm tentatively supports the former explanation, as some of the duck bones found at this site look similar to those of wild species, though the possible presence of very small domestic birds cannot be discarded. DNA analysis, which has successfully discriminated between bones of wild and domestic geese at the Saxon site of Flixborough (Barnes et al. 1998; 2000; Haynes 2000), may be able to provide an answer.
Although only one duck bone (an ulna from Period 3 - Early Roman) bears butchery marks, it is likely that most (if not all) duck and goose bones derive from birds that were eaten. The absence of ?rock dove bones (Columba livia/C. oenas) at Heybridge is surprising, considering that they have been found in many Roman sites in Britain (Parker 1988). Domestic doves are known to have been raised in the Roman period, but it is not yet possible to securely distinguish the bones of the domestic dove/pigeon from those of the wild form (Columba livia) and the osteological distinction between rock and stock dove (Columba oenas) is similarly problematic (though see new criteria in Tomek and Bocheński 2009).
The largest wild bird found is the mute swan, identified through a piece of sternum from the early Roman period (Period 3) (Table 118). Distinction of mute (Cygnus olor) and whooper (Cygnus cygnus) swan bones is difficult as the two species overlap in size, but the sternum of the whooper swan has a very distinctive hole in the anterior end of the keel. The fragment from Elms Farm was identified as mute swan as this feature was absent. Swan bones were also found at Colchester (Luff 1993) and at six other sites in central England. In two of these - Caister-on-Sea (Norfolk; Harman 1993) and Wroxeter (Shropshire; Armour-Chelu 1997) - the presence of mute swan is claimed. Parker (1988) mentions other Roman sites in Britain where bones of the other swan species were found.
As mentioned above, some duck wild species are so small that they cannot be confused with domestic forms. This is the case of the teal (Anas crecca), found at Elms Farm in the sieved material from the early to mid-Roman and Late Roman-Early Saxon periods. This small duck is recorded from nine other sites in central England, including the nearby Colchester (Luff 1993), and has also been found in several other localities in the rest of the country (Parker 1988).
The exploitation of water environments is confirmed by the presence of bones of several waders, such as grey plover (Pluvialis squatarola) and curlew (Numenius arquata) (Table 118 and Table 119). These birds could have been caught in the estuarine environments that abound in the vicinity of Heybridge. Shore birds occur quite regularly in large Roman assemblages, including at Colchester (Luff 1993), where both (golden) plover and curlew were found. Parker (1988, 204) suggests that these birds could have been 'caught by fowlers and brought in for sale at market'.
Woodland species are represented by the woodcock (Scolopax rusticola), which is one of the most common birds found in Roman sites. This species has also been recorded at Colchester (Luff 1993) and Caesoromagnus (Luff 1992) and was presumably hunted for its meat.
An interesting bird, frequently recorded for Roman sites in Britain, is the raven (Corvus corax), found in small numbers in most periods at Elms Farm too. Although the possibility these birds were eaten cannot be excluded, their scavenging habits could have made them rather unappealing as food. Among the many possible reasons for the common presence of ravens in Roman sites, Parker (1988) mentions the possibility that they could have been kept as pets - apparently they keep well in captivity - with the added bonus that they could have scared hawks away. He also suggests that they may have had a symbolic meaning. Ravens are nowadays almost exclusively present in the west of the country (Snow and Perrins 1998, 1485), but their geographic distribution in the past may have been different.
Another noteworthy occurrence at Elms Farm is that of the peregrine falcon (Falco peregrinus), represented by a single bone found in Period 4 (mid-Roman). As far as we can ascertain, this is the first record of this species in a Roman site in Britain. The presence of the peregrine, alongside that of other birds of prey such as gyrfalcon (Falco rusticolus), merlin (Falco columbarius), goshwak (Accipiter gentilis) and sparrowhawk (Accipiter nisus), would, in a medieval site, be interpreted as possible evidence of hawking. However, falconry is not known to have been practised by the Romans and it is therefore not surprising that such species are rarely found in Roman sites. None of them tend to scavenge and, therefore, if not used by people, they are unlikely to end up in an anthropogenically formed assemblage. Scavenging raptors, such as red kites (Milvus milvus) and white-tailed eagles (Haliaeetus albicilla), are much more common (see Parker 1988). The finding of a sparrowhawk bone in association with many thrush bones - the most common prey of the trained sparrowhawk - at another Essex site, Great Holts Farm, has led to the suggestion that in localised cases the Romans may have used raptors to catch other birds (Albarella 1997; Murphy et al. 2000). It is questionable whether in these cases an activity similar to falconry can be proposed or other techniques, such as the use of the bird as a decoy, should be regarded as more likely. The accidental occurrence of peregrine bones in an archaeological assemblage is unlikely, but cannot be entirely ruled out.
The large size and well-preserved nature of the bone assemblage from Elms Farm has resulted in the collection of a considerable amount of information about many aspects of the site. This information has been useful at many levels, from helping to understand the nature of various areas within the site and the local economy to how the process of Romanisation affected the site, and how the site fits into the regional and wider context. Most of these issues have been addressed above in relation to specific pieces of evidence, so this discussion aims to bring these individual elements together to present a more coherent summary of the evidence from the site as a whole.
As at most Iron Age and Roman sites, the major domestic species, cattle, sheep and pigs, dominate the assemblage from Elms Farm. This was true for both the NISP and MNI values. The relative frequency of these three main species did differ between the two quantification methods and when the sieved material was assessed separately from the hand-collected bones. The relative abundance of the sheep seems to be under-represented in both the NISP counts and the hand-collected material. This is to be expected, given the smaller size of sheep bones. While cattle completely dominate the assemblage in the later periods, sheep form a larger proportion in the earliest periods.
It is interesting to note that while the major domesticates and many other species normally found on Roman sites were present at Heybridge, one conspicuous exception was noted. This was the absence of domestic cat, which, while never common on Roman sites, is usually represented by a few bones within a large assemblage. The absence of cat bones is not a recovery issue as similarly sized bones of small dogs, hare and badger were recorded, as well as many smaller bones from birds and small mammals. It may be that cats were only owned by people above a certain status, as very few have been found on more rural sites. Most cat bones have come from major urban centres such as York (O'Connor 1988), Lincoln (Dobney et al. c.1996), Colchester (Luff 1993) and Exeter (Maltby 1979), exceptions being the villa at Great Holts Farm (Albarella 1997) and the religious complex at Ivy Chimneys (Luff 1999).
One interesting feature of the site as a whole is the way the settlement patterns within the site shifted through time while maintaining the focus on the public spaces. This movement is seen in the changes to the buildings within different areas of the site through time (Atkinson and Preston 1998) and also the patterns of rubbish disposal. It seems that in each period the area used for the disposal of vertebrate remains displays less activity in terms of buildings and occupation evidence.
During this period cattle bones were most prevalent but a significant proportion of sheep bones were also present. Pig was present in all periods with a roughly constant, if small, frequency. Slight spatial differences were noted in Area K, with a more equal split between cattle and sheep and a higher percentage of pig bones. A few chicken bones were recovered. Most animal bones were found in Areas H and L and to lesser extent in D and K. Evidence of butchery on species other than cattle was limited in all periods and in this period cattle butchery was much less intensive than in the later periods. The age-at-death data show that cattle were mostly killed when they had reached the subadult category while the sheep and pigs were split between the subadult and adult categories. The incidence of non-metric traits on the cattle mandibles was low, while for the sheep a very high frequency of premolar foramina was noted. The size of cattle and sheep in Period 2 was quite small and was similar to most other Iron Age sites. Sheep were about the same height as the group of Shetland ewes used as the standard, but slightly more slender limbed. Pigs were also small, slightly smaller than Neolithic pigs used as the standard. Horses were generally quite small, except for a few large individuals, and fit well within the range of Iron Age horse from the rest of Britain.
As in Period 2, cattle provided the largest number of bones but again a significant proportion of sheep were present. This was particularly noticeable in Area J where the concentration of sheep was high (see discussion of the temple area below). Additionally, in Area I the proportions of cattle and sheep were equal. Area I was the main area used for disposal of animal remains in Period 3, the only period in which this area was extensively used for this purpose. The intensive butchery of the cattle bones appears in deposits from Period 3 onwards, as do the specialised butchery practices that produce the characteristic hook-damaged scapulae and 'soup-kitchen' deposits. The mortality profiles of cattle and pigs remains very similar to the preceding period but the age at death of the sheep is slightly younger, apart from those associated with the temple complex (see discussion below). Several non-metric traits show different frequencies in this period; the proportion of cattle M3s with reduced hypoconulids fell but the number of mandibles with abnormal mental foramina rose. The proportion of sheep mandibles with either premolar or abnormal mental foramina fell. A major change occurred during Period 3 in the size of the cattle, which increased significantly. However, all the other animals stayed much the same size as in Period 2, apart from a few large individuals among the sheep and horses.
There was a change in the species proportions in this period, as cattle dominate the assemblage by a much larger margin than before. The exceptions were in Areas D and I where cattle and sheep are roughly equal. Disposal of rubbish was changed again in this period, and was mainly concentrated in Areas H, J and K. Changes were also noted in the mortality profiles, with cattle mostly killed as adults rather than subadults. More elderly individuals were also present. The sheep pattern was similar to Period 2, with most animals killed as subadults. The pig mortality profile showed that although most were killed as subadults, there was a shift to the younger end of the range with more immature rather than adult individuals present. The non-metric traits show some changes, with the percentage of cattle and sheep mandibles with abnormal mental foramina increasing again. The biometry shows major size increases in sheep, pigs, horses and chickens, whilst cattle size does not change from Period 3 (i.e. they were still large).
As in Period 4, cattle bones dominate the assemblage by a large margin. Rubbish disposal seems to have been confined mostly to Area J, in a series of pits encroaching on the edge of temple precinct. The age-at-death data for all three domesticates is similar to that in Period 4. The relative frequency of non-metric traits changes again, including a marked increase in the number of cattle mandibles with abnormal mental foramina. There was also a slight increase in the frequency of M3s with a reduced hypoconulid and a decrease in absent P2s. The sheep mandibles show a decrease in the frequency of both abnormal mental and premolar foramina. The biometry shows no change in animal size.
Although there was less material from the final period, cattle bones still dominate the assemblage but there is a slight increase in the number of sheep. Most rubbish disposal continued to be in the pits encroaching on the edge of the temple precinct (Area J). The pattern of mortality profiles is similar to the preceding two periods. The number of cattle mandibles with abnormal mental foramina falls but the relative frequency of other non-metric traits remains roughly the same. The size of the animals at Heybridge in this period continues to be as large as those in the preceding two periods, which is unusual as other sites in Britain are showing a decrease in size at around this time (Dobney et al. c1996). The horses are a possible exception to this, as they show a visual but not statistically significant decrease in size.
The size and age variations mentioned in the section above are summarised in Tables 150 and 151. These clearly show that the greatest period of change occurred between the Early and mid-Roman periods.
| Cattle | Sheep/goat | Pig | Horse | Chicken | |
|---|---|---|---|---|---|
| Late Iron Age/Early Roman to Early Roman | increase | no change | no change | no change | ? |
| Early Roman to mid-Roman | no change | increase | increase | increase | increase |
| Mid-Roman to Late Roman | no change | no change | no change | no change | |
| Late Roman to latest Roman/Early Saxon | no change | no change | no change | slight decrease | no change |
| Cattle | Sheep/goat | Pig | |
|---|---|---|---|
| Late Iron Age/Early Roman to Early Roman | no change | decrease | no change |
| Early Roman to mid-Roman | increase | increase | slight decrease |
| Mid Roman to Late Roman | no change | no change | no change |
| Late Roman to latest Roman/Early Saxon | no change | no change | no change |
One of the most important research questions at this site was the detection of the transition from the Iron Age to the Roman period. This is a particularly difficult question to address, as these changes need not be all that visible/evident. This may be due in part to the fact that the pre-Roman Iron Age societies in this part of Britain were fairly sophisticated in their organisation and economy (Wacher 1998). Therefore the changes required to become 'Roman' were not as great as in other parts of Britain. In addition, the transition is more likely to have been a slow process of the accumulation of 'Roman' attributes, rather than the introduction of a 'package deal' at one point in time. At Heybridge it appears that a mixture of these two routes to Romanisation took place.
At Heybridge there appears to have been two 'waves' of Romanisation that affected the economy and animal husbandry. The first happened very soon after the conquest and in the zooarchaeological evidence showed up in the form of a size increase in cattle, the introduction of particularly Roman butchery practices and changes in the frequencies of several non-metric traits. The underlying animal husbandry regime appears to have remained as it was in the pre-Roman periods. Millett (1990, 11) complains that the dearth of late pre-Roman Iron Age evidence prevents us from understanding whether large breeds of animals were introduced to Britain in that period or as a consequence of the Roman arrival. The availability of a complete chronological sequence at Elms Farm allows us to suggest that this phenomenon was triggered by the arrival of the Roman invaders. As the south-east of England (particularly Kent and Essex) was the area first occupied by the Romans following the invasion of Britain (Wacher 1998), it is unsurprising that Heybridge acquired some Roman characteristics during this period. The physical appearance of the settlement, which had already undergone significant transformation prior to the conquest, was submitted to further change (Atkinson and Preston 1998) suggesting that although it was a relatively minor settlement it was important enough to the invasion force to be worth bothering with.
This may reflect the fact that the Roman army's first settlement point was Colchester, where they stayed to await the arrival of the Emperor before embarking on more campaigns. During this period there would have been a need to establish supply mechanisms to feed the army. Heybridge is very near to a navigable river, which would make it useful as a settlement on a supply route. The encampment of the army at Colchester and Camulodunum (Millett 1990, 59) would have meant that the surrounding area was required to provision the troops. As the pre-Roman agrarian economy in this area included a substantial proportion of cattle already, the provisioning of the army with beef would not have demanded many changes to the husbandry regime. The size increase shows that larger cattle were rapidly introduced to the area to improve the local stock. Whittaker suggests that the army would be provisioned with its own supply, independently from the 'vagaries of the market' (1989, 69). Since some of the food items would have been obtained through long-distance trade it is possible that some of these larger cattle were introduced by the army itself. Millett (1990, 56), however, disagrees and states that the army preferred to use local supplies and that, in any case, the food demands of the occupation were modest, the army forming no more than 2% of the total population. If we accept Millett's model it should consequently follow that the improvement in local livestock must have been imposed more by an acculturation concern than by the needs of the invading army.
The second group of changes were more widespread and occurred around a century after the conquest took place. These comprised size increases in the rest of the domestic species, more changes in the frequencies of non-metric traits and shifts in the mortality patterns of cattle and pigs; cattle to reflect the kill-off of older animals and pigs the opposite pattern. The changes in the age-at-death of the domestic species suggest that the underlying economic basis for keeping the animals had changed and/or that cultural factors had altered, i.e. the classic description of Romanisation.
In the case of cattle it is most likely that the economic emphasis had shifted. It was expected that a shift towards younger animals would have taken place, as the demand for beef grew. However, at Heybridge it appears that older animals were favoured. This suggests that either dairy products became a more important resource or that the animals were primarily used for traction and secondarily as a meat source. In view of the high incidence of pathological conditions that can be caused by traction and the absence of neonatal and juvenile cattle, the latter explanation is more plausible. In addition, Romans are not known to have been keen on cows' milk (White 1970), although we do not know to what extent this could be applied to areas outside the Mediterranean. Intensification of cereal agriculture would have meant that many more draught oxen were required, both for the agricultural process and transport.
The greater number of juvenile pigs may be a consequence of husbandry intensification and cultural taste. Very young (suckling) pig was considered a delicacy in Roman times (Varro II.4.19) and the book of recipes by Apicius mentions pork far more frequently than any other meat. These written sources may only refer to high-status meals and to an earlier period, but the importance of pork in the Roman diet in Italy is unquestionable and is supported by much zooarchaeological evidence (e.g. King 1978; 1984; 1985; 1994; 1999). There is a hint that the diet of the inhabitants of Elms Farm was becoming more influenced by Roman taste.
This shift in the economic base and diet of the inhabitants and the increased size of all livestock suggests that the transition to a Romanised existence was a more gradual process than the initial changes to the physical appearance of the settlement and the provisioning of the army. The site illustrates the point that while the outwards appearance of the settlement was essentially Roman from immediately after, and to some extent even before, the conquest, the agricultural practices of the inhabitants and those in the surrounding countryside maintained some Iron Age characteristics for a further century. A possible explanation for why these changes took place in the middle of the 2nd century is the fact that around this time the army had to be reinforced to quell rebellions in Wales, northern England and Scotland (Wacher 1998). This may have had the extra effect of increasing the rate of Romanisation around this time because of the increased 'Roman' population (i.e. ex-soldiers) throughout the country. As King (1984) has pointed out, the 'Roman' soldiers did not need to come directly from Italy, but were likely to have been recruited from other provinces in central Europe. Consequently the diet of the Roman army in Britain may have been influenced by a number of different traditions, ranging from Italy to Germany and Britain itself.
This two-stage Romanisation of the settlement at Heybridge means that there is no clear boundary between the Iron Age and the Roman periods, with the two merging as a continuous process. In addition, the final result is not truly Roman in the sense that the two cultures have merged, rather than one being replaced by the other, hence Romano-British rather than Roman. Also the animal economy is very different from that seen in Italy and the Mediterranean heartland of the Empire.
Placing the animal bone assemblage from Heybridge in the wider context is a complex task, owing to the diversity of Romano-British settlements with animal bone assemblages that have been excavated and published. Deciding which sites to use for comparison was not too difficult, as for many the bone assemblage was too small to be useful. Within the local area a large number of sites have been dug, particularly in the towns (Colchester, Chelmsford and Braintree), and several rural sites (Harlow, Ivy Chimneys, Great Holts Farm) have also yielded useful assemblages.
During the later Iron Age the Elms Farm assemblage is similar to most other sites in the south and east of Britain. The only slight anomaly relates to the high proportion of cattle, which is at least partly the result of recovery bias but is also consistent with sites in the East of England at this time (Hambleton 1999). The size of the animals at Heybridge is also consistent with data from other Iron Age sites.
The animal bones from the Romano-British periods present a more complex picture. There is only one site near Heybridge (Braintree) that seems to have the same kind of status (i.e. a secondary urban centre or small town), which has made wider comparisons a little difficult. The major urban centres in Essex at Colchester (Luff 1993) and Chelmsford (Caesaromagus) (Luff 1982) would have had a very different economic base to small towns, which were essentially rural in character. Most animals would have been brought into the major urban centres on the hoof from a very wide catchment area and many different populations. This means that there is great potential for bones from a very diverse range of animal types, sizes and ages to be found in urban deposits, whereas at Heybridge the bones are more likely to have come from animal populations in the local area in which there would have been less variation.
In spite of these difficulties a few points are worth making about the comparisons between Heybridge, the major urban centres and some rural sites. At Colchester and Chelmsford the predominance of cattle in the assemblage is very marked in the later periods but, as at Heybridge, the sheep in the earliest periods form a significant proportion of livestock. The age profiles from both Colchester and Chelmsford show a predominance of younger individuals in all three main species indicating a meat-based economy, as would be expected in an urban setting. There are a few changes in the later periods with older sheep and younger pigs being noted, which were also observed at Heybridge. At both urban centres there was a steady change in the size of the main domestic species through time rather than a substantial change at one point in time as seen at Heybridge. This is most likely the result of the animals coming from several populations that may not have been improved at the same time or to the same extent. The major urban centres in Essex are reflecting what is happening in the more rural towns such as Heybridge, but with a time delay as the changes filter through.
The one small town in Essex that has a useful zooarchaeological report is Braintree (Smoothy 1993). The assemblage was smaller than Elms Farm but still of a size sufficient for meaningful analysis. One limitation on comparison with Heybridge is the fact that there was very little material from Early Roman deposits so the analysis concentrated on material from the mid-2nd century AD onwards. Species proportions appear to have been very similar, with a high percentage of cattle followed by sheep, pig, horse, dog and deer. Butchery patterns on the cattle bones from Braintree seem to be similarly intensive to those at Heybridge, which represents a further indication of the adoption of a 'standard' Roman butchery practice. Cattle and to a lesser extent sheep, seem to have been mostly mature when killed whereas pigs show a marked peak of younger individuals. The measurements of the domestic animals (all small quantities) fit within the data from Chelmsford and Colchester. From this it seems that the economy at Braintree was based on the same principles as that at Heybridge and the animals in the mid- to Late Roman period were of a similar size.
Many of the rural sites excavated in Essex appear to have a specific function. Two sites, Harlow (Legge and Dorrington 1985) and Ivy Chimneys (Luff 1999), were dominated by religious complexes, with very little in the way of associated settlement, indicating that the bone assemblages are dominated by the activities of the temple. Hence the different species proportions are a reflection of this rather than the economy of that area. The similarities and differences between these temples and that at Heybridge are discussed in a separate section below.
Similar problems are encountered when trying to compare villa sites with other types of settlement. Firstly, the term villa has been applied to a variety of rather different sites, the only linking theme being the fact that these are all isolated settlements mostly presumed to be lived in by an extended family rather than a community. The differences in status between the many settlements described as villas are very obvious in the type and quality of the buildings and the interpretations of the nature of the settlement are also very varied. Most do, however, appear to be some kind of farmstead. This means that they are the producer sites from which the smaller and larger urban centres are supplied, and will differ from them in that respect. One villa of this type is at Great Holts Farm (Albarella 1997) where, although the assemblage is small, a predominance of cattle is evident. In addition, the presence of some very large cattle may indicate that, as a primary producer, the site received imported cattle that were used to improve the local stock. Cattle from predominantly consumer sites such as towns and, to some extent, Heybridge itself, were probably smaller as a result of cross-breeding.
The similarities in the zooarchaeological evidence between the two small towns of Braintree and Heybridge are striking, as are the differences between them and the major urban centres at Colchester and Chelmsford. This suggests that the delineation between major urban centres and small towns (or secondary urban centres) can also be defined in terms of the zooarchaeological evidence.
Beyond the south-east of England there is a sharp decline in the number of Roman small towns and rural sites in Britain that have been excavated in Britain. This may reflect a real decrease in the density of settlements but it is perhaps more likely to be the result of the unevenness of the archaeological research, which itself partly derives from differential development in different areas (including in this case the more intensive recent development of the south-east of England. Consequently the zooarchaeological evidence for much of the rest of Britain comes from other major urban centres where modern development has resulted in the excavation of Roman deposits. These include Exeter (Maltby 1979), Lincoln (Dobney et al. c1996) and York (O'Connor 1988). All these cities were founded initially as military institutions that became civilian settlements at a later date. In this respect they are similar to Colchester. The excavations at Lincoln and Exeter were within the centres of the Roman settlement while at York the excavation was within the vicus rather than the military town itself. In many respects the zooarchaeological evidence reflects this, with Lincoln and Exeter being more similar to each other than to York.
In the early military periods at Exeter and Lincoln the age profiles show an emphasis on younger animals, which is hardly surprising given that these are most likely to be the remains of meat provisions for the army. At York, and at Lincoln in the later periods, sheep in particular and cattle to a lesser extent mirror the older animals seen at Heybridge. Perhaps civilian market forces demanded secondary products and good quality/prime meat was a commodity not directly catered for. Similar butchery patterns were seen on cattle bones from all these sites and were particularly noticeable at Lincoln. The size of the animals was only compared with Lincoln as the data were sparse or not easily available for the other two. Both cattle and pigs were substantially smaller than those at Heybridge. At Carlisle (quoted in Dobney et al. c1996) the cattle were even smaller. This suggests that the influence of imported stock to improve local animals may have become less and less noticeable the further north the site location.
Lauwerier (1988) undertook an extensive study of Roman and native settlements in the Dutch eastern river area, which provides a very useful summary of data with which to compare British sites. In terms of straight distance, Holland is closer to sites in Essex than those in the far north of Britain so the comparisons are still useful. The species proportions are significantly different between most of the Dutch sites and those in Britain. While the percentage of cattle remains is high (60-91%), the proportions of sheep and pigs on Dutch sites are the reverse of those seen on British sites with pig providing a greater proportion of bones. This is particularly true of the military sites. It is likely that this is a product of differences in both the environments of the sites and the underlying Iron Age husbandry regimes. In Britain sheep husbandry was more widespread in pre-Roman times, whereas the eastern river area was renowned for its pig production. In addition, the areas surrounding some of the Dutch sites are more suitable for rearing pigs than sheep.
In her study of bone assemblages from the north-western provinces Luff (1982) also picked up on the fact that the suitability of the surrounding environment for either pig or sheep rearing was reflected in the animal bone assemblages. In particular it was noted in the north German plain area (Todd 1975) and also in areas of France. The proportion of cattle was dependent on both the type of site and the surrounding environment. Military sites were generally cattle dominated while at civilian sites environmental factors played a greater role.
The age profiles of the assemblages from Holland were, on the whole, very similar to those seen at Heybridge and Braintree. Around 50% of cattle lived beyond four years of age. Lauwerier suggests that meat production took second place to the importance of cattle as draught animals as has been hypothesised here for Heybridge. The Dutch sheep were also killed at an older age than would be expected from a primarily meat-based economy, again indicating that secondary products were a valued resource. Interestingly, pig bones indicated that while most of the pigs were killed before skeletal maturity, there was more of a spread of ages than at Heybridge. White (1970) mentions that there is no evidence that livestock were deliberately bred for eating in the Roman period, a statement that seems to be backed up by the evidence of cattle and perhaps sheep but not pigs, which provide little in the way of secondary products.
Very similar butchery patterns - in particular the hooked scapulae and the 'soup kitchen' deposits - were also noted on cattle bones from Dutch sites (Lauwerier 1988), from the famous site of Augusta Raurica in Switzerland (Schmid 1972) and from other north-western Roman provinces (Luff 1982). This shows that Roman butchery techniques appear to have been adopted across a very wide area.
In terms of the biometry, the Dutch cattle were larger in early and late periods than at Heybridge but about the same in the mid-Roman period. This suggests that in Holland the principles of stock improvement were carried further throughout the Roman period while in Britain the initial size increase was maintained rather than improved upon. A size increase is evident in the other animals around a century after the conquest of Britain, this appears to have happened a century earlier in the dutch area. This indicates that the improvement of animals was a slow accumulated process in Holland as in Britain. This size increase is seen across the north-western provinces with the remains of larger sheep being recovered from Roman sites, although generally the dating is not refined enough to detect when this occurred. In general Dutch horses were slightly larger than their British counterparts in all periods.
The site of Elms Farm contains an area that appears to be related to religious activities, which continued in use from the Iron Age through the transition and right through the Roman period. Its existence is not surprising as many pre-Roman religious centres in Britain were located on tribal boundaries and were often associated with trade centres for the simple reason that in a sacred place quarrels were less likely to break out between traders (Wacher 1998). This fits with the hypothesised market area at Elms Farm and the settlement may well be on or near the boundary between the Trinovantes and Catuvellauni tribes.
The fact that the pre-Roman temple at Elms Farm was rebuilt in the same place immediately after the conquest is quite a normal phenomenon. The same thing happened on many sites across the Empire and particularly in Britain and Gaul as part of the Romanisation process (Millett 1995). Although the Romans discouraged worship of non-Roman deities, they did not want to alienate the conquered populace completely so they fused the native deities with existing Roman gods or gave them more Roman names and rebuilt the temples in a more Roman manner (Goodman 1997). As no evidence has been found to link the temple cella at Elms Farm with any of the major deities of the Roman pantheon, perhaps one of these Romano-Celtic hybrid deities was the focus of worship here. Concentric temples (as seen in the Heybridge complex during Period 3 (Atkinson and Preston 1998)) are usually known as Romano-Celtic temples and are distributed across the parts of Britain and Gaul that were occupied by pre-Roman Belgic tribes. Small circular Romano-British temples (seen in all periods in the Heybridge complex) are also known and are usually a continuation of existing structures (Wacher 1998).
As there was no evidence to link the Heybridge temple complex with a specific deity and hence a known set of rituals, the evidence from the animal remains and other finds has a useful potential in providing some insight into the activities that took place there. During excavation a few pits were tentatively identified as related to the activities of the temple complex, mostly because of their proximity to the temple buildings rather than the finds located within them. A reasonable quantity of animal bone was recovered from the sacred area around the temple buildings. However, much of it was recovered from a series of rubbish pits, which encroached upon the temple precinct in the later Roman periods (Periods 4-5 onwards), and are therefore unlikely to be associated with the activities of the temple itself. Although none of the contexts stood out as containing obviously ritual material, more detailed analysis has given a few pointers to some of the activities at this temple.
Most of this centres on the concentrations of chicken bones and an age at death profile from the sheep mandibles, which differed from that seen across the rest of the site. Taking the chicken bones first; 70% (sixty-two) of these were recovered from the temple precinct and most of these (fifty bones representing a minimum of seven birds) were recovered from two Period 4 pits centrally placed within the sacred area. While the numbers of bones involved is quite small, the imbalance in the distribution is striking and is unlikely to result from differential preservation as the preservation across the site is relatively uniform. This suggests that these concentrations of chicken bones may well be the result of activities associated with the temple. Assemblages of chicken bones are known from a number of sites defined as ritual/religious. Seven headless chicken skeletons were also found in association with the temple at Caesaromagnus. The lack of the head was, however, attributed to a taphonomic bias rather than to ritual processes (Luff 1992). The remains of a cockerel were found in a post-hole at the Romano-British site of Wavendon, near Milton Keynes, and were thought to represent a ritual deposition (Williams et al. 1995). Chickens are known, from written sources, to have had a ceremonial function in the Roman world (Toynbee 1996, 257), and archaeological evidence for their possible use in religious activities has also been reported from outside Britain. Cremated bones of selected parts of chicken carcasses are reported from 1st-century AD levels from the House of Amarantus at Pompeii (Italy). Powell (1995-6, 104) suggests that they may represent remains of birds used for cock-fighting or in sacrifices Although the Elms Farm chicken bones associated with the temple are not burnt, a remarkable similarity is represented by the association between chicken and sheep bones, which was also noted at Pompeii. In addition, as at Elms Farm (see above), male individuals predominate at Pompeii. Toynbee (1996, 257) specifies that cocks in particular were sacrificed to the Lares (household gods). Findings from the northern Italian site of Montereale Valcellina indicate that such practices may date back to the Iron Age. Evidence for the use of chickens, sheep and pigs was found in ceremonial pits of this early Romanised settlement (2nd-1st century BC). The closest parallels to these activities are to be found in Celtic necropolises of the north-eastern Italian region of Veneto (Petrucci and Vitri 1995).
Looking at the sheep mandibles, it was observed that the mortality profile was different in Period 3 (Figure 638). The suggestion was put forward that as the temple complex was most active during this period, perhaps this was influencing the kill-off pattern and Figure 639 confirms this hypothesis. It was expected that young animals would be associated with the temple, as at Harlow (Legge and Dorrington 1985) and Caesaromagus (temple site; Luff 1992), however, all the other areas show high numbers of young animals and the temple precinct has a high number of older adult animals. The Romano-Celtic temple site at Ivy Chimneys, Witham (Luff 1999) also had a large number of older sheep associated with it.
Perhaps the reason for this was that young animals were too precious a commodity to sacrifice and that animals past their 'sell-by-date' were used instead. This is borne out by the fact that at Chelmsford (mansio site; Luff 1982) a large number of the sheep remains from the temple area were pathological. Indeed the practice of using weak and sick animals as sacrifices is mentioned in Varro (II.v.11). Alternatively, animals in their prime (possibly male, as females were of greater economic importance) may have been the preferred sacrificial offering at this temple. This seems to have been the case at Uley (Levitan 1993) where a large number of male goats were found, and also at Chelmsford (Luff 1982) where several rams were identified from the horncores.
The mandibles from older individuals were all located in two pit complexes (dating to Period 3), one directly in front of the temple cella to the side of its entrance and the other directly behind the temple building. Because of the proximity of these pits (and those containing the chicken bones) to the temple building it has been suggested that they were probably associated with temple activities, even though no 'votive deposits' were identified within them at the time of excavation (Atkinson and Preston 1998). It should, however, be remembered that these small quantities of possible sacrificial material from Heybridge were found in pits that contained much the same range of domestic type rubbish as was found in pits across the rest of the site.
Another aspect of religious activity on the site appears to be associated with some of the wells. At Heybridge, eleven wells were excavated and their contents analysed. These wells were spread across the site and represent all periods of activity. The animal bones from a few of these wells appear to differ from those in other wells and from the fills of pits and other features. Foe example, most obvious of these is the fact that most of the articulated whole and part skeletons were recovered from only two features, a deep pit and a well (three contexts, see Table 120). The skeletons recovered from the Heybridge wells included those of cattle, horse, dog and juvenile pig. It is a frequent phenomenon on archaeological sites of all periods that articulated remains are found in wells. It is often presumed that these animals were put down the wells as a convenient place to dump them and that this was after the wells fell out of use. However, there is also the possibility that these animals represent votive offerings.
Cunliffe (quoted in Luff 1982) stated that a 'surprisingly number' of deities worshipped in the Roman World were associated with water. Many sites across northern Europe have revealed 'votive' deposits placed in watery places, usually bogs, lakes and rivers or wells. In Britain there is a particular concentration of these deposits in the south-east (Luff 1982). Most of these deposits seem to occur in the Late Iron Age and Roman phases of occupation (although earlier and later examples are known). Dogs, horses and juvenile animals are particularly commonly found in deposits of this type and many also include isolated skulls. Feature 6641 (Group 579, Area H, Period 5) was originally interpreted as a well but is now thought to be a purpose-dug deep pit. Pit 6641 contained a reasonably complete horse skeleton but the head (both cranium and mandibles) was missing. This is slightly unusual as most such deposits were the other way round; perhaps the head was displayed near the temple complex as a reminder of the sacrifice, while the carcass was used as an offering. This pit also contained five pewter bowls. Metalwork was also often associated with wet contexts (Atkinson, pers. comm.). The practice of sacrificing animals in a well is described by Horace in his Odes (III, 13 quoted in Luff 1982) and seems to imply that this was done while the well was still in use. This is corroborated by the fact that many of these skulls and skeletons are found right at the bottom of wells. The pit in Area H is near the temple precinct and is also in a public space, which is perhaps why it was used as the location for this putative 'votive' offering.
The second feature (Well 14984, Group 710, Fill 20034, Period 4-5) contained the remains of a complete cow, at least four whole dogs of various sizes, two juvenile and one neonatal pig and a few juvenile sheep/goat bones. This well is in Area L, some distance from the sacred area. Dogs were often associated with healing and death in the Romano-Celtic world (Luff 1982) and are often found in similar situations to horses. This may not necessarily have anything to do with religious activity but may relate to the disposal of animals that were not normally eaten. Juvenile animals are also often associated with votive deposits but may equally be the result of natural casualties as hypothesised for pigs (see above). The deposits in this well date from the Late Roman period when changes were occurring at the site and it is suspected that this is when the well fell out of use and was filled up. The horse in pit 6641 is considered more likely to be a votive offering because of its location in a public place and the headless nature of the skeleton. However, the possibility that this is just the disposal of an unwanted carcass cannot entirely be ruled out.
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Terms and Conditions | Legal Statements | Privacy Policy | Cookies Policy | Citing Internet Archaeology
Internet Archaeology content is preserved for the long term with the Archaeology Data Service. Help sustain and support open access publication by donating to our Open Access Archaeology Fund.