A North-Western Habitat: the Paleoethology and Colonisation of a European Peninsula
Rogan D.S. Jenkinson
Cite this as: Jenkinson, R.D.S. 2023 A North-Western Habitat: the Paleoethology and Colonisation of a European Peninsula (a comprehensive analysis of excavations in Pin Hole Cave, Creswell Crags), Internet Archaeology 61 (Monograph 31). https://doi.org/10.11141/ia.61.1
6. Temporal Survival, Reduction and Inter Species Relationships for the Major Vertebrate Species and Genera
The intention of this section is to describe and summarise the large collection of vertebrates known from Pin Hole Cave, as follows:
Accurately identify species and genera housed in current collections and especially to incorporate the total vertebrate faunal evidence rather than previous selected elements of the faunal population.
Reconstructing where possible their original location within the cave and its deposits.
Describe remains in terms of their survival, based upon minimum number of individuals and bone fragments.
Describe the variation in body parts within the cave in an attempt to gain an insight into their Quaternary history.
Describe the morphological and surface damage to bone fragments to assist in the interpretation of taphonomic events within the cave.
6.2 Large carnivores
6.2.1 Spotted Hyaena (Crocuta crocuta)
The single species, Spotted Hyaena, is well known from European Quaternary localities. For modern species, individual body length varies between 1.2-1.6m and height to the shoulder is usually between 76cm to 91cm, and they can achieve a maximum speed of 65km per hour (Walker et al. 1975). The species is nocturnal and often lives in caves or dense vegetation. Individuals live in packs and show a large degree of cooperation in hunting and scavenging. Females are polyestrous all year and average 2 to 3 young, and are known to move the location of litters to avoid predators. Males lack seasonal sexual activity, (Walker et al. 1975). The species is equipped with a powerful dentition and mandibular muscles, which are able to break and crush vertebrate bone of much larger species. Studies by Klein and Scott (1989) of size variation in Hyaena premolars from a number of fossil localities suggest that the occupants of Pin Hole Cave are some of the largest recorded. The species is one of the most versatile and powerful of predators. Studies by Kruuk (1972) have shown that although they are commonly thought of as scavengers, both sexes cooperate in hunting and obtain most food requirements in this way. These studies have also demonstrated that the species caches food resources, and those sites of residence are often littered with the remains of prey, a practice that has earned them a reputation as 'bone accumulators'. Excavations of a modern Hyaena den by Sutcliffe (1970) have noted that many collected bones are gnawed and that bone with spongy shaft interiors are often licked clean, an action that produced a distinct polish and striations on the bone surface. Much of this activity is due to juvenile behaviour where young individuals regard the bones as 'toys'. These studies have also recorded that adult individuals have the ability to carry relatively large weights or body parts, such as heads and part carcasses (Kruuk 1972).
Vertebrate bone from such localities is therefore distinct in appearance with spongy bone chewed at both proximal and distal ends, producing scratch striations or gnaw marks often accompanied by surface polish. The presence of bone in this condition may well indicate residence and denning, as opposed to occasional sheltering, behaviour. In his 1970 study, Sutcliffe recognised four types of damage to bone; splintering, juvenile gnawing and scalloping, producing hollow bone shafts with striations and polishing, and digestive damage producing bone flakes with scalloped surfaces and holes.
Packs of Hyaena have few modern competitors and are quite able to defend their kill sites against other large predators such as lions. Many of the Creswell localities, including Pin Hole, have long been recognized as Hyaena dens, associated with large quantities of damaged vertebrate bone. Many studies have interpreted the species' presence and activity as the prime taphonomic factor and few attempts have been made to examine the temporal and spatial evidence that would support such a claim.
The collection of bone held in UK museums from Creswell derived from this species numbers 784 fragments. This figure includes remains recovered by Mello, Armstrong and the passage clearance by the author. There are 386 specimens documented by Armstrong as two-dimensional records on each object. These are incorporated within the reconstruction. In general terms, the reconstruction is based upon 48% of the recorded items in two dimensions. Addition of the illustrated passage remains to this general area within the reconstruction would mean that the total reconstruction represents 68% of the recorded remains.
There are three main concentrations of this species, which are:
The area within 63-66ft (18.9-19.8m) north from the entrance and within the depth 5-12ft (1.5-3.6m) below the stalagmite datum. Within this concentration the remains are within the mound of sediment accumulated under the roof rupture and they are also inclined down a southern facing slope. A total of 30% of the illustrated remains occur within this area.
The area within 50-62ft (15-18.6m) foot north of the entrance and within 5-12ft (1.5-3.6m) below the stalagmite datum. Remains within this area occur within the lower area of sloping mound sediments. They consist of 20% of the illustrated remains.
The third concentration is within the area from the cave entrance 14-50 ft north (4.2-15m) and within depth levels 4/5-10ft (1.2/1.4-3m) below the datum. Within this area the remains appear to have a more horizontal distribution.
The distribution suggests that the main concentration of Hyaena occurrence is within the cone of sediment immediately underneath the roof rupture in the area to the rear of the cave. The area of their distribution diminishes toward the entrance. The frequency of juvenile and foetal individuals is fairly consistent within all three concentrations and averages between 13%-17% of the illustrated population. Despite the observation that there are more remains within the cave rear, the ratio of juveniles present suggest breeding and denning activity throughout the cave and within a reasonably stable circumstance over a long time period.
The uppermost boundary of the concentration coincides with the occurrence of the slab layer recorded by Armstrong, which is now seen as evidence of the upper surface of a lag deposit. This suggests that there is a hiatus of unknown duration between the rear concentration and those remains known from the front of the cave.
The upper distribution between 20-55ft (6-16.7m) north and 1-5ft (0.3-1.5m) depth is distinct. The origin of this material is not obvious. This does not appear to show disturbance of sediment and deposits. This is particularly indicated by the consistency of distribution of the remains, which themselves concentrate within stratigraphic levels 4-5. The relative variation of body parts represented is practically the same as those known from the cave rear, which also seems to indicate that the cluster is not random and the result of postmortem movement. The huge concentration of (visible) vertebrate bone within this cave has already been mentioned and as a result it is possible that some of this material has been utilised by later human groups and incorporated within deposits where their remains are frequent. The reconstruction highlights the difficulties of interpretation caused by the inability to reconstruct in three dimensions. Examples of vertebrate bone distribution that are the result of carnivore activity are usually distinct. A comparative example (see Taphonomy of Recent Vertebrate Skeletal Remains report) shows the debris associated with a Wolf denning site in Jordan where bone debris is seen strewn around the entrance area in a random fashion, clearly showing the presence of semi-articulated body parts and although gnawed bone is present, it is found on a number of bone fragments.
Post-mortem movement - human or carnivore curiosity?
Within the upper areas of distribution for Hyaena, there are a number of unusual occurrences of remains that are difficult to explain or interpret. Many comments have been made in previous studies concerning individual specimens, particularly where one piece is broken and found in different locations. This is an important aspect of the evidence that has been used to suggest disturbance. The more spectacular occurrences are:
Area 66/2ft - a complete mandible of an old Hyaena. Both sides of the mandible are fused together and are fairly heavy. The specimen was probably on a slope of sediment but at 2ft (0.6m) from the surface datum of stalagmite. It is very difficult to imagine an object of this size and weight sliding or rolling (from where?) within the sediment at this location and this area is one of the higher areas of infill. Its presence is an enigma. One possibility is that it has been moved by a human occupant of the cave during the Later Palaeolithic when the only carnivores within the cave were humans and European Wolf. Circumstantial evidence for possible human interest exists in that the upper levels contain Upper Palaeolithic lithics as well as a Belemnite fossil, a Jurassic Echinoid from Lincolnshire, an amber fragment, a piece of mother of pearl from a marine mollusc and a decorative marine shell, most of which were discovered within this general area of the cave.
Areas 54/5ft and 54/6ft (16.2-1.5/1.8m) contained another complete adult mandible. The location is probably a southern sloping deposit and the items are separated by 0.3m depth from the datum but within levels perceived as of similar age (and providing evidence of an incline within the sediment within a central area of the cave).
Area entrance 14-50ft (4.2-15m) within depth 1-4ft (0.3-1.2m). This area contains 18 specimens of Hyaena, mainly small teeth, which occur in stratigraphic levels normally believed to be coeval with the Later Palaeolithic. Most of the specimens occur in level 3 (depth 0.9m) and seem to be deposited horizontally. This is an interesting phenomenon and could be post-mortem movement of older remains into sediments of younger age. This is not a significant factor and does not undermine the stratigraphic credibility of the surrounding sediment. The context of this evidence is that the 'derived' remains are 0.025% of the vertebrate bone content of the cave, which seem to have moved southwards by up to 10m.
Hyaena Population survival
The Hyaena remains are represented by large numbers of bone fragments found throughout the cave sediments. A total of 784 bone fragments of an estimated 61 individuals have been included in this study. Indirect evidence is available in the form of coprolites, not included here but which are reported from several areas of the cave. A complete record of remains are summarised in the digital archive and in spreadsheets by stratigraphic level.
Figure 22: Hyaena (Crocuta crocuta) temporal distribution and survival (source data: Digital archive)
The analysis is based specifically upon fragments of the species recovered from the excavations of Armstrong from 1924 onwards, and these have been grouped into stratigraphic levels according to his dimensional records.
The survival of bone remains for the species is shown in Figure 22, which expresses the surviving percentage of bone in respect to that expected within a live population. This has been calculated on the minimum number of individuals present in each stratigraphic level. It clearly demonstrates that the remains are derived from a relatively low number of individual animals. There is considerable variation in the survival of bone fragments of this species. The most frequent occurrence is within the lower levels, where MNI estimates are between 10-15 individuals while the upper levels average between 5-10 individuals
The lower stratigraphic levels, particularly Levels 11 and 12 contain a significant concentration of skeletal remains that are dominated by juvenile and foetal individuals. These are particularly concentrated in the cave rear at the terminus of the passage and adjacent to the surviving sediment section. The reconstruction of the species' survival suggest that there are three distinct temporal concentrations, between levels 12-13, levels 8-10 and levels 3-5.Many of the bone remains are derived from the cave rear, an area where there is a distinct slope in sediments down and towards the cave mouth. In these circumstances the horizontal reconstruction suggests that the highest distribution within the stratigraphic sequence is contemporary with the Late Quaternary (Upper Palaeolithic artefacts), yet this is unrealistic in that the Hyaena distribution is mainly from the upper parts of the slope and the human evidence is concentrated on the slope itself. Presence of the species is is also available in the form of distinctive bite and gnawing damage, which is present not only on their own bone remnants but also on a range of other species in the form of both bite and gnaw marks.
In general terms, bone survival averages less that 10% of a potential live population for most of the sequence and with a distinct decline in both individuals and bone within the upper parts of the sequence. The apparent very low rate of survival may have multiple causes but is likely to be the result of both the general decline in frequency of the species through time and their known modern-day behaviour, where they consume and destroy their own species. The high frequency of survival for parts of a low number of individuals within the lower part of the sequence may be due to residence within the cave, or rapid burial resulting from a geomorphic event. The low incidence of individuals may indicate occasional, seasonal use of the site and most probably by small family-based groups.
Figure 23: Well-preserved crania of Hyaena (Crocuta crocuta) from stratigraphic level 7. Reproduced courtesy of Manchester Museum
Despite the high frequency of bone destruction and survival, many bones are well-preserved. Figure 23 illustrates one of several well-preserved Hyaena skulls complete with dentition known from the sequence. Complete preservation of such skulls is relatively rare in such sediments as the Hyaena themselves would often destroy them.
In summary, the pattern of distribution shown in Figure 22 appears to indicate an initial population that has declined in numbers of individuals over time. The relatively dense population characterising the early levels (12 to 15) is represented by an erratic distribution in younger levels, representing a 30-40% decrease in individual numbers accompanied by a relatively stable frequency of individual survival and preservation, all of which seems to indicate a dramatic reduction of Hyaena activity within the site. The stratigraphic levels where the species decrease in numbers coincide with those that indicate human use of the cave. This contrast may indicate that human groups are responsible for such decreases in the species presence at a time when Hyaena populations appear to be in general decline. Many studies have suggested that the species disappeared from the UK some 30,000 years ago and before the Last Glacial maximum. There are indications of stratigraphic distribution within this cave that suggest that the species may have survived in later periods. Clear evidence of this possibility is made difficult by the massive concentration of vertebrate bone within such a relatively small area and despite being recorded fairly well, there is little contextual information.
Figure 24: A complete juvenile skeleton of Hyaena (Crocuta crocuta) discovered during the excavations by Jenkinson
The recovery of a juvenile skeleton occurred during modern excavations during 1986, at a depth of 4.35m, in an area that correlates with the reconstructed excavation sequence of Armstrong of 12ft (3.6m) (Stratigraphic level 12). The complete skeleton is well-preserved and is a very rare occurrence for such a fragile vertebrate body (see Figure 24). Its presence must indicate not only residence for breeding but perhaps rapid burial, which has ensured its survival. Many juvenile remains of this species are also preserved as fragments within the cave sequence. The presence of well-preserved remains of juvenile individuals suggests that the locality has been used as a breeding den by the species. This contrasts quite markedly with evidence of older Hyaena individuals from this cave and from other Creswell sites where the species is present and juvenile bones are rare.
The notable preservation of foetal and skeletal parts from young individuals is graphically illustrated by the near complete preservation of foetal mandibles and post-cranial skeletal parts from the rear of the cave (Figure 25, Figure 26). This is in marked contrast to the heavily damaged adult mandibles also found within the cave rear.
Figure 25: Hyaena (Crocuta crocuta) foetal mandibles from the cave rear
The distinct lack of damage to mandibles suggests that the remains were either rapidly buried in collapsing den areas or that the breeding location was abandoned by adults. The semi-articulated individual discovered during the 1984 excavations suggests that rapid burial either caused death or immediately followed it.
Spotted Hyaena attrition
Figure 27: Survival of Hyaena body parts (source data: Digital archive).
Large numbers of surviving Hyaena teeth survive, particularly within the earliest levels, which are followed by a dramatic reduction in frequency within level 10 and 11. In the stratigraphic levels above these, teeth survival increases within level 9 and then gradually decreases throughout the upper sequence. The frequent survival of teeth as fossil remains of this species has been observed at other UK Quaternary locations (Turner 1981). Frequency of other body parts is significantly lower within the whole sequence and is dominated by limb parts, where axial remains are confined to level 10. For limb bones the low frequencies known from levels 11 to 13, are followed by a 40-50% reduction for the whole of the upper sequence. The frequency distribution of body parts, particularly those for dentition, agrees in temporal aspects with that of recorded species survival and also suggests greater presence of the species within the lowest levels, suggesting residence or denning within the cave. While limb part distribution follows the general trend, their frequency is not in proportion with dentition survival and suggest that these parts either represent a lower number of individuals present or that they have undergone more extensive reduction. In these circumstances the more resilient teeth are more likely to survive.
Hyaena bone damage
Fragmentation of vertebrate bone by Hyaena has been observed on both modern and fossil sites. The damage is often very distinctive and indicated not only by teeth marks but also more commonly by morphological change in bone shape. Figure 28 illustrates such reduction by gnawing and biting in examples of Hyaena mandible (see also digital archive) and Woolly Rhinoceros humerus (Figure 29). The progressing degree of destruction is very clear, and is corroborated by the presence of Hyaena teeth marks. The reduction develops in a distinctive pattern, which is irrespective of species variation.
Many of the remains of this species are well-preserved but have been heavily damaged, particularly by fracturing and gnawing by other individuals of the same species. See selection of examples in digital archive. Many skeletal parts have additional surface polish resulting from licking by juveniles (e.g. V154).
Although there is considerable diversity in the survival of respective skeletal parts, the remains are dominated by the survival of teeth. In this instance, it seems that other parts of the maxillary and mandibles have been fractured or gnawed to destruction or at least into small fragments that cannot be identified to this species (e.g. V18).
Figure 28: The general character of Hyaena (Crocuta crocuta) bone attrition indicated by a fossilised Woolly Rhinoceros humerus
Similar patterns of bone damage are evident from the cave, particularly on the skeletal parts of large ungulate species. Lightly gnawed Reindeer antler, which are often from cast pieces, is also fairly common. Damage type on these skeletal parts have not been systematically recorded in this study but the nature of the damage seems to vary. In the basal and older levels fracturing, while destructive, does not appear to extensively destroy the bone. These antler pieces often have the distinctive gnawing marks of Hyaena teeth. In contrast the upper levels have similar but less destroyed antler with lighter gnawing damage with fewer and smaller teeth marks, which is likely to indicate damage by Wolf rather than Hyaena. Bone damage is particularly distinctive in some larger species, including those of Woolly Rhinoceros. There are multiple examples of distinctive damage by Hyaena which include:
etching and perforation of bone flakes by Hyaena stomach acids - see V4
coprolites - often containing small bone fragments
Figure 29: Humerus of Woolly Rhinoceros (Coleondonta antiquatatis). A typical example of a Hyaena-damaged limb bone. The proximal has clear remnants of bite marks along the edge and the distal medial and lateral surfaces have clear marks showing that the bite penetrated the entire thickness of the bone. The shaft retains a highly polished surface (under the varnish), most likely the result of surface licking probably by juvenile Hyaena. (Image courtesy of Manchester Museum)
The variation of attrition for bone remains also indicates similar temporal change that was noted for body part survival (see Figure 30). Attrition type varies considerably through time. In general terms, peak frequency for both complete and fractured bone remains exists within the lower levels. In these levels, the relative frequency of fractured bone coupled with a low occurrence of chewed and gnawed bone suggest a different form of activity by the species.
Figure 30: Hyaena (Crocuta crocuta). Attrition, bone damage (source data: Digital archive)
A second but lower peak of occurrence occurs within levels 7-9 and is associated with a smaller incidence of complete bone and gnawing. This pattern is reproduced on a smaller scale within levels 3-5. The data for attrition types suggest three distinct phases of activity by Hyaena, levels 11-13, levels 7-9 and levels 3-5, and each of these are clearly separated by evidence of a much lower indication of attrition activity. All of the evidence is within a context where the level of activity declines temporally. A single fossil skull of Hyaena, originally from the passage, has very uncommon damage to the crania where the nasoid area is fractured and missing (e.g. V165). There is no sign of gnawing anywhere on the skull and the multiple fracture surfaces and edges do not look modern. This may be an example of damage from movement or rock fall within the sediment.
There is also a decline in frequency of bone flakes, those that cannot be identified to species, on the basis of their morphology. In addition to their distinctive shape and form, many pieces are penetrated by variable size holes and the surfaces have a scalloped appearance and sheen or polish. It was originally suggested by Armstrong that one particular piece (Figure 31) might be a 'Bull Roarer', similar to those known from anthropomorphic studies of Australian aborigines (Armstrong 1936). It was suggested that the hole within the right-hand side of the bone was to attach a thong, which would allow rotation to produce a sound. It is now known that this is one example of a number of such pieces from the sediments, and which are the remains of partially digested vertebrate bone that have been regurgitated by Hyaena (Sutcliffe 1970). As such, these are direct indicators of Hyaena presence within the cave. It is known from modern studies (Sutcliffe 1970) that Hyaena have preferred locations or spots for regurgitation of both bone flakes and hair. The nature of the excavation recording system makes identification of such places within the cave very difficult, but it can be demonstrated that many within the early sequence were discovered within the cave passage.
Figure 31: A vertebrate bone flake showing evidence of partial digestion, including scalloped surface and holes and which has probably been regurgitated by Hyaena (Crocuta crocuta), Pin Hole Cave (Image courtesy of Manchester Museum)
Stratigraphic distribution of vertebrate bone flakes has traditionally been associated with Hyaena predatory activity (Mello 1876; Campbell 1977; Jenkinson 1984). The temporal and spatial distribution of bone flakes has been reconstructed and is illustrated in the digital archive. The reconstruction is based upon the re-plotting of all the flakes that have sufficient dimensional information to record their original stratigraphic location prior to excavation.
The flake distribution clearly shows that although they are frequent, the stratigraphic distribution is clumped and not continuous. They also do not closely coincide with the stratigraphic distribution of Hyaena. Within level 12, a large number of flakes occur that may have been associated with residence by Hyaena. Within stratigraphic levels above this, the flakes, while numerous, are not frequent within the overall assemblage and show a tendency to increase in younger levels. This is associated with a low and decreasing occurrence of gnawed bones (see attrition flakes distribution). Within some of the youngest levels, their occurrence is associated with a very low occurrence or absence of Hyaena and a significant rise in the Wolf population. This suggests that the lower sequence has Hyaena gnawing activity on bones of large ungulate species, associated with residence by the species, which in the upper sequence is replaced by Wolf and associated gnawing, and most probably of different smaller species. There is no obvious correlation between this activity by Hyaena and levels where there is evidence of human use of the cave, with the exception of levels 11 and 12, which have evidence of cut-marks on both Woolly Rhinoceros and Mammoth. In contrast, flake occurrence probably resulting from Wolf activity is associated with both the youngest evidence of the Middle Palaeolithic and levels with Upper Palaeolithic lithic assemblages. This observation is in close agreement with the result of bone damage by fracturing, where significant numbers of fractured bones are known from levels with archaeological evidence but is accompanied by very low frequencies of gnawed vertebrate bone belonging to Wolf.
Summary
The evidence recorded for Hyaena presence within the cave suggests that:
From an early peak of use there is a long-term decline in the species presence and activity
The combination of evidence and the clear presence of juvenile and gnawing suggest use of the site as a den for breeding and where at least one juvenile individual has survived in a complete state,
Three distinct episodes of residence are suggested by the separate occurrence of attrition and gnawing and where the most recent two episodes are on a relatively reduced scale.
The distinct clumping of temporal distribution may be explained in several ways and could be the result of environmental change.
It is also possible that human use of the cave has disturbed long-term seasonal use by Hyaena, particularly as the archaeological evidence reflecting human activity is concentrated within and around the low incidences of Hyaena between the peak frequencies. It has been suggest that Hyaena and human groups may have had a sympatric relationship (White and Pettitt 2011), where they may have scavenged from each other's kills. There seems to be very little supportive data for this view. The stratigraphic distribution of the species and its overall reduction in frequency during the Pleistocene contrasts with evidence for human presence and also shows an interrupted or clumped temporal presence. In addition, there is clear evidence of presence of Hyaena activity, which includes breeding, denning and juvenile chewing and gnawing of vertebrate bone. Evidence for human activity is different, with cut-marks on bone, albeit of similar species but the known examples have no evidence of carnivore damage. The overall indication suggests that use of the site for each group is not contemporary. The low incidence of Hyaena activity within the upper sequence may reflect the longer-term trend toward localised extinction of Hyaena during the Quaternary and which has been recorded at several English sites. A causal factor in this process is that there is evidence of interspecies competition between human groups and Hyaena populations at this site where competition would have been of significance.
6.2.2 European Wolf (Canis lupus)
Modern and recent fossil populations of this animal are grouped in the single species Canis lupus, but are variously referred to as the Timber Wolf, Grey Wolf or European Wolf. Adults are often between 1-1.4m in length and just less than 1m in height, and with an average weight of 25kg. (Walker et al. 1975; van Den Brink 1977). The species is an intelligent and social creature and congregate in social groups or packs of between 10 to 20 in number, and cooperate in hunting.
Diet is varied and is composed of smaller species such as rodents, fish and carrion, which has been graphically described by Mowat for Arctic populations (1963). In some circumstances, wolf packs are known to specialise in group predation of Reindeer. Individuals are known from modern studies to fracture, chew, gnaw and swallow the bones of prey species. Breeding often utilises dens or natural shelters such as caves and takes place between January and March. These localities are usually abandoned when the newborn are around 2 months of age. Individuals can often travel at between 40-50km per hour.
A total of 382 bone remains of European Wolf (Canis lupus), estimated to be derived from 33 individuals, are known from Armstrong's excavation. Of these, 104 have sufficient dimensional details to incorporate them in the reconstruction. A further 37 bones can be located to the passage lower levels. The reconstruction is therefore based upon 36% of the known remains.
Individual remains are grouped by body part for cranial, axial and limb. There are two separate concentrations of European Wolf remains. The first is in the cave rear, in the area between 57-67ft (17-20m) north from the entrance and in stratigraphic levels 6-10ft (1.8-3m). The form of this concentration suggest that it accumulated on a south-facing slope. The second concentration is within the area 33-66ft (9.9-19.8m) and within the upper stratigraphic levels 1-4ft (0.30-1.2m) north from the entrance. The form of the distribution suggests that the depositional environment is horizontal.
The main lateral concentration of European Wolf remains occur between 44-71ft (13.2m-21.3m) from the cave entrance and between depths 2-19ft (O.6-3m). Remains for the lower levels are recorded as a group that occur between 58-78ft. A cluster of remains in the rear of the cave between stratigraphic level 8-12ft (2.4-3.6m) for each lower level and reflects an assemblage of remains occurring along the slope of sediment below the roof rupture. Remains above this stratigraphic area accumulated along slightly more horizontal sediments for the length of the cave.
European Wolf Population survival
The osteological remains are described and summarised for each stratigraphic level in the digital archive. An analysis of this species' survival (Figure 32) shows that the population increases during the Quaternary. There are three distinct and 'sudden' increases in levels 9-13, levels 6-7 and in the uppermost levels between levels 2-5. Their skeletal remains, when compared to those of a live population estimated by MNI, show a high degree of destruction, averaging 90% of expected parts. Minimum number of individual estimations (MNI) indicate that most levels contain the remains of less than 5 individuals and in ten of the levels only a single individual is present. It has been estimated that levels 5-12 accumulated over a 50,000 year period. They contain 195 wolf skeletal parts from 16 individuals, which in practical terms means that at least one wolf was in the region every 2000 years.
Figure 32: European Wolf (Canis lupus). Temporal distribution and survival (source data: Digital archive)
There is a clear similarity in their temporal distribution to that of human use of the cave (Figure 7). The clustering of their distribution is remarkably similar to that shown in Figure 7 where Middle Palaeolithic artefacts occur between levels 10-12 and 6-9, with Upper Palaeolithic artefacts in levels 1-3. The low incidence of their occurrence suggests that the cave was not used extensively for residence or breeding. Despite the richness of the fossil record for this species, it is impossible to distinguish events or detailed associations between remains. Despite the timescale, the long-term association in the distribution of humans and wolves is intriguing
Wolf attrition
The surviving body parts of this species, preserved as fossils, are dominated by head parts, associated with a lower number of limb parts, with axial or central body parts practically non-existent (Figure 33).
Figure 33: European Wolf (Canis lupus) attrition, body part selection (source data: Digital archive)
The distinct discrepancy between the survival of head parts with other elements of the skeleton and the close correlation between these and the distribution of human occupation may indicate a selective use of wolves by human groups (such as skinning). The dramatic survival of head parts within levels 5-12 seems to be unrelated to their resilience, and contrasts with the upper levels of 1-4 that show an increase in survival of all body parts.
Figure 34: European Wolf (Canis lupus), attrition, bone damage (source data: Digital archive)
Damage to wolf bone is illustrated in Figure 34. This clearly shows that many of the specimens are fractured parts of the original bone. It is also clear that carnivores have gnawed very low numbers of bone fragments, including the wolves themselves. The fragility of the remains of this species and their consequent destruction by large carnivores, such as Hyaena, the slight rise in numbers and the large rise in bone survival later in the cave's history when the large carnivore population is reduced or extinct, occurs at a time when human groups are present within the cave. This suggests a relationship between the two species. The dominance of complete and fractured bone with a low incidence of gnawing in levels where few other carnivores are present suggests that the Wolf population inflicted the damage, a factor that would suggest occupation by the species.
The temporal distribution of vertebrate bone flakes and gnawed flakes within the uppermost part of the sequence is associated with wolves in levels where Hyaena is absent. Much of the flake material cannot be firmly identified to species from their morphology but their general size suggests that they are largely derived from small ungulates such as Deer, presumably Reindeer or Bovids.
Analysis of the remains of Hyaena and European Wolf clearly shows initial residence by Hyaena groups associated at least with Woolly Rhinoceros, its probable main prey. The bones of both species have evidence of massive destruction by Hyaena. The frequency of this species declines dramatically in level 7 in the closing stages of the Middle Palaeolithic. Following the species decline, there is a very dramatic rise in the presence of European Wolf and where the population appears to be largely an adult one with little direct evidence of young. This dramatic change is associated with levels containing extensive evidence of the Upper Palaeolithic. Carnivore damaged and gnawed vertebrate bone is associated with both species and there appears to be no clear method of distinguishing the respective culprits. There are several examples (Figure 35) that show extensive carnivore damage by Hyaena or European Wolf.
Figure 35a-b: European Wolf skull and mandible with evidence of fracturing and gnawing. Originally recovered by Armstrong but not reported by him. They are described for the first time in this study.
A skull sample has been extensively gnawed and is reduced to a fragment of left maxillary (Figure 35) and a mandible has been gnawed around the ramus leaving the fore segment of jaw, and which illustrates typical Hyaena gnawing (cubs?).
Although not quantified, there appears to be a greater frequency of stomach-acid damaged, regurgitated bone of larger species in levels associated with Hyaena. The implications of the stratigraphic overlap of Hyaena and Middle Palaeolithic evidence and separately of Wolf presence with evidence of the Upper Palaeolithic are difficult to assess. The presence of Hyaena young individuals is not indicative of any particular season (Kruuk 1972). During such a period, both Hyaena and Wolf groups would have made use of a number of similar locations for developing young, as a method of predation avoidance. It is inconceivable that the species are coeval.
Wolf presence within the cave is of two types. The lowest stratigraphic levels (5-12) have very low numbers of individuals, whose remains survived primarily as fractured head parts associated with very few post cranial bones, which tend to be phalanges and metapodials (i.e. feet bones). This body part can easily be swallowed and destroyed by larger carnivores such as Hyaena. This strongly suggests that both Wolf and Hyaena used the cave at different times. There is distinct survival of heads and feet, some of which are slightly damaged, particularly in a context with little if any carnivore gnawing. This may be good evidence for the exploitation of Wolf as a source of pelts by Neanderthal groups. The evidence for the presence of both species temporally vary but correlate closely.
In the upper levels of the cave (stratigraphic levels 0-4), wolves are more plentiful and represented by all body parts and notably by axial parts and a large decrease in head parts. In these levels they are associated with extensive human use of the cave. This raises the possibility that at this time, the two groups developed a symbiotic relationship. There is an associated presence with both species of cast (collected) Reindeer antler which has not only been cut and worked but has been gnawed as well, sometimes all on the same Reindeer antler! This also very strongly suggests that species habitation was coeval and that the relationship was developed possibly to the extent that both humans and individual or small numbers of wolves were not only living together but may have been cooperating in their daily lives during the Upper Palaeolithic.
6.2.3 European Lion (Panthera Leo)
This species seems to have enjoyed an extensive distribution within varied habitats during the recent past. Descriptions of Quaternary populations rely heavily upon studies of modern-day populations, particularly those studies in the Serengeti and Kruger National Parks (Schaller 1976). Modern day individuals have a body length between 1.8-2.4m and a body weight of between 181-227kg (Walker et al. 1975). The species is polygamous and breeds throughout the year with an average litter of 2-3 individuals, although larger numbers are known. The species prefers open aspect country such as plains. Hunting can be by single individuals or in groups or prides, particularly of females, and a preference is shown for large ungulate species. The dentition is adapted for meat eating and they have highly manipulative paws, used to bring down prey species. At kill sites, lions are known to gorge on the remains of most of their prey, and occasionally leave some axial skeletal parts. The remnants of prey can vary depending upon interspecies competition with other carnivore species (Kruuk 1972). They are not normally known from caves.
A total of 46% of the European Lion remains have been incorporated into the reconstruction of distribution and can be located to the passage area. The species is represented by a low number of bones and these occur in the area over 50ft (15.2m) north from the entrance. They are randomly distributed within this area. Their occurrence is unusual in that they are represented by cranial, limb and axial body parts. This suggests that whole or part carcasses may have been within the cave. European Lion may have been occasionally resident within the cave.
European Lion Population Survival
A total of 146 vertebrate bones, estimated to be derived from 15 individuals, were identified to this species. The temporal distribution of 14 individuals is shown in Figure 36. The species is predominantly present in stratigraphic levels 8 to 14 with a less frequent occurrence within levels 1 to 4. The population is predominantly adult but two juvenile individuals are also present. Bone survival within the cave, in comparison with that of a live population, is very low and there are often less than two individuals in any particular level.
Figure 36: European Lion (Panthera leo) temporal distribution and survival (source data: Digital archive)
Lion attrition
All parts of head, limbs and axial bones are preserved in low numbers and their variation is illustrated in Figure 37 (see survival and reduction, reduction Lion body part, attrition). A larger number of head parts occur in stratigraphic levels 12 and 10. The analysis indicates that, more or less, Lions are represented by near-complete carcasses within the cave and that these particularly occur in levels that have evidence of Neanderthal lithics and Hyaena activity.
Figure 37: European Lion (Panthera Leo) body part selection
The distribution of body parts shown in Figure 37 is specific and interrupted. In general terms, stratigraphic levels 7 to 13 have a discrete population. Levels 1 to four also contain a low number of individuals. In general terms, reduction and the surviving bone from the live population is extreme with an average of between 25-5% of individuals represented. In addition, the few surviving remains are dominated by head parts with a few limbs, many of which are tarsals and carpals.
Bone damage is illustrated in Figure 38, which shows that most skeletal parts are fractured and these are associated with a few complete bones, mostly metapodials. There is an exceptionally low incidence of gnawing by carnivores and Hyaena. The unusual lack of substantial attrition and gnawing contrasts with attrition of other occurrences of megafaunas within the lower stratigraphic levels.
Figure 38: European Lion (Panthera Leo) attrition, bone damage (source data: Digital archive)
The lack of damage to skeletal parts is illustrated in Figure 38. The relative survival of heads and metapodials associated with fewer but damaged limb and axial parts may indicate skinning of pelts from carcasses. The virtual lack of gnawing associated with these remains indicated an absence of Hyaena and supports the observation that this indicated human activity.
Figure 39: Right mandible of European Lion (Panthera Leo) from stratigraphic level 10 in the rear of the cave (Image courtesy of Manchester Museum)
The distinct and well-preserved right mandible of a juvenile Lion (Panthera leo) was excavated within level 10 (Figure 39). The mandibular surface is unfused, indicating the mandible is from a younger animal. This mandible is the one held by Leslie Armstrong in a photograph taken in the cave entrance. A small area of damage on the ramus has truncated the bone. There is insufficient morphological evidence to suggest that the fracture is natural or carnivore induced. The remaining surface of the bone bears no trace of carnivore gnawing or bite marks. The medial and mid-surface of the bone bears six elongate and oblique marks. All of the marks have lightly broken the bone surface and have been mineral-filled following deposition. The damage is difficult to interpret in that the marks are clearly not modern and appear to reflect an attempt to separate the tongue from the mandible. The surface damage is probably the result of lightly-executed cutting actions. The medial surface additionally bears traces of surface polish, which is particularly concentrated around the epiphyseal gap. Studies by Kitching (1963) suggest that this may have been a 'ripping tool'. The extensive medial surface polish is typical of surface licking by juvenile Hyaena but may also have been the result of use. The presence of surface cutting marks, surface polish and the lack of Hyaena gnaw marks in a stratigraphic context where surface gnawing of large vertebrate bone is frequent, suggest that this mandible is an artefact. Kitching's original suggestion that is a 'ripping tool' is a possibility or, conversely, it may simply be a waste product from Neanderthal skinning activity.
6.2.4 European Brown Bear (Ursus arctos)
Despite great variety in both modern and Quaternary types within Northern Europe, studies by Erdbrink (1953) have grouped them all into the species Ursus arctos. The species displays considerable variation in size, diet and behaviour. In Europe, individuals are usually about 2m in length and vary between 150 to 750kg in weight (Walker et al. 1975). They are sexually dimorphic and are powerful species, largely immune from predation (Kurtén 1958; Ewer 1973). The species normally hibernates during winter, when the young are born, in natural shelters including caves. Diet is omnivorous but can vary in different habitats. Predation, where present, is often concentrated on the slower moving ungulates, especially Bison, but the main diet consists of plants and occasionally fish. Individual bears are known to occupy life territories of up to 75km².
European Brown Bear is represented by 307 bone fragments, and 86 (26%) of these have sufficient dimensional details from the excavation to allow reconstruction of their distribution within the deposits. The remains of bear occur throughout the cave length and depth. There is a significant cluster of remains within the passage and rear of the cave. As is the case with other carnivores, these seem to be concentrated within the mound of sediment underneath the roof rupture. In front of this feature, remains are randomly distributed within the upper sequence from the entrance to the rear slope.
European Brown Bear population survival
Osteological remains of this species are itemised and summarised for each stratigraphic Level. Fragments occur in very low individual numbers within most levels (Figure 40). In most cases, bone survival relative to that of a live population is reasonably high. There are a number of milk teeth among the surviving bones which indicate the presence of females and foetal or newborn individuals. There are many modern records of females adults not having sufficient nourishment for young during partial hibernation, which results in the death of both. At least one male individual has a fractured baculam which has healed in life. The presence of Bear suggests an absence of other larger carnivores.
Figure 40: European Brown Bear (Ursus arctos) temporal distribution and survival (source data: Digital archive)
European Brown Bear survival is very variable and ranges from 43% to 22% within stratigraphic level 10 (Figure 40). Levels 1, 3-4, 5-6 and 12 all have 0-10% survival of bone from 11 individuals. An exception is layer 12 where nearly a quarter of the individual remains survive. There is an intriguing occurrence of European Brown Bear within the area of 64ft (19.2m) north from the entrance and in the 2ft (0.6m) depth level where parts of a single individual is represented by four metatarsals, all of which display the development of bony tissue resulting from osteo-arthritis.
European Brown Bear attrition
The species is represented by a wide variety of body parts, which are shown in Figure 41. These are dominated by the frequency of head parts but associated with both limb and axial parts. Body part survival is consistently similar and proportional to the variation in the number of individual animals known for each layer.
Figure 41: European Brown Bear (Ursus arctos), body part selection (source data: Digital archive)
Survival of a wide range of body parts suggests that the remains are derived from natural casualties, perhaps dying during hibernation. The survival of juvenile bones suggest females and newborn dying during hibernation in a context where no or few other predators are present and where most of single individuals are incorporated within the sediment. There are a number of milk teeth from the cave which also indicate hibernation and winter occupation. Corroborative evidence exists in the nature of bone attrition, where high numbers of bone are complete and associated with simply fractured bone. The presence of gnawed and carnivore damaged material probably indicates scavenging of decomposing carcasses by carnivores, particularly Hyaena, during the summer.
Figure 42: European Brown Bear (Ursus arctos), attrition bone damage (source data: Digital archive)
Relatively large numbers of Brown Bear skeletal parts survive in a complete or simply fractured form and the incidence of carnivore gnawing is low in frequency (Figure 42). Gnawing is present in the lower stratigraphic levels of the cave and is clearly associated with Hyaena, whose frequency within the deposits diminishes and finally disappears over time. Gnawing of Brown Bear bone in higher levels of the cave, when Hyaena was absent, must have been the result of Wolves. The dramatic increase in species numbers and remains within the upper levels of the cave may indicate a change in circumstances - an increase in the frequency of this species and the relative lack of large carnivores.
European Brown Bear (Ursus arctos) skull
Within the population of Brown Bear, a single skull from layer 10 or 11 is exceptionally well-preserved and of exceptional size (Figure 43). The individual is from alarge full-grown adult male and is complete but has broken zygomatic bones and damage to both parietals. There is no evidence of carnivore damage to the skull. The central region of this skull bears distinctive and unusual damage. In the mid cranial region, not only are both zygomatic arches broken with sharp breaks, but also the orbital cavity behind each side has extensive but both parietals have shattered and fragments have been lodged inside the crania.
Figure 43: European Brown Bear (Ursus arctos) skull from stratigraphic level 10 or 11 (Image courtesy of Manchester Museum)
When examined (in 1976), neither cavity had been excavated but superficial examination suggests that the source of fracture is on the left side of the skull and an impact here has pushed fragments into the crania, preserving them. The right side of the skull has a large area with similar fresh breakage but the fractured pieces appear to be missing and are not apparently preserved within the infilling matrix. The skull has not been examined by removing the sediment. The examination relies solely on the external morphology. The unusual nature of the breakage in an otherwise well-preserved skull is of interest and suggests that the fractures were caused by an impact on the left side of the skull probably from the rear, and which has fractured the zygomatic arch and entered the orbital, passing through the parietal. The same action appears to have exited on the right side, shattering the parietal and orbital and separating these fragments from the main skull. Access to these areas of skull for the object that inflicted this damage must have been at near horizontal to the long axis of the skull and has produced fractures in the same region on both opposing sides of the skull. The overall nature of the breakage and its specific localised nature do not seem compatible with rock fall fracture, particularly as the damage is located in the restricted orbital sockets area. There is also no evidence of carnivore damage in the form of biting or gnawing and it is also difficult to envisage such damage given only the zygomatic bones and recessed orbitals are damaged. Although the evidence is circumstantial, the nature of the break suggests that a reasonably narrow projectile has been driven from the left side of the skull and into the crania and that this has broken through the right side of the crania. There is no surface morphological evidence that would indicate the timing of such an event. It is also unlikely that the damage could have been the effect of a postmortem event within the sediment build up or rock fall within the cave. The nature of such a wound would almost certainly result in death and given the size and potential danger of the animal concerned, this may well have happened during hibernation in the cave or during a summer visitation by human groups.
6.3 Summary: large carnivore occurrence
The stratigraphic distribution of large carnivore species is summarised in respect of bone frequency within Figure 44. Long-term trends in species' occurrence throughout the Quaternary are distinct. Hyaena populations show a variable but gradual decline in number and consistently occur with the remains of European Wolf. Although Wolf occurs with less frequency, it appears to slowly replace Hyaena and exhibits a significant increase during the latter stages of the sequence. Brown Bear is consistently present throughout the whole sequence but does display a significant increase in mid-sequence.
The existence of inter species competition within such a carnivore population presents a number of problems for the interpretation of their palaeoecology and particularly as the species are very unlikely be have been coeval. An additional difficulty is that in some cases the occurrence appears to coincide within concentrations of human occupation. Despite the uncertainties presented by the excavation and lack of clear distinction of stratigraphic levels, it seems possible to draw a number of clear conclusions.
Figure 44: Temporal distributions of large carnivore species (source data: Digital archive)
Hyaena populations dominate within the mid- and lower sequences. The presence of juveniles, bone fragmentation and bone surface polish indicate annual residence for breeding. Residence could have taken place at any time of year but the presence of fragmented and gnawed bone of large ungulate species and juveniles indicate a summer or late summer presence where the locality was utilised for short-term denning of young. The overall trend of occurrence diminishes over time and there is little indication why. It is unlikely that inter-species competition could have caused such a decline, although there is evidence in the form of cut and separately gnawed bone that suggests human and carnivore occupation was coeval in the sense that both may have utilised the locality at different times in any given annual season.
The consistent and continual presence of European Wolf and the increase in occurrence during the later stages of the sequence is intriguing. Its occurrence, seemingly, varies and increases during human occupation and is associated with similar variation in the occurrence of its potential prey in the form of Reindeer and sudden increases in a wide range of both vole and bird species. The presence of juveniles and the stratigraphic occurrence with gnawed vertebrate bone (where Hyaena are of low frequency or absent) suggest use of the cave for breeding. The low incidence and regularity of occurrence, notably at times of increased human occupation of the cave and the presence of gnawed and cast Reindeer antler (of no potential food value), may suggest coeval use of the cave with human groups and the existence of a symbiotic relationship between the two groups.
European Brown Bear has a distinct occurrence in low frequencies seemingly within the cave passage and rear. In at least one case, the extensive parts of one individual are preserved within the cave rear in association with the remains of Grayling scales. Large numbers of bone fragments lack evidence of gnawing by carnivores and there is some possible evidence of the death of one individual from injuries to the skull. The most likely explanation for the species' occurrence is that the remains are from the death of hibernating individuals throughout most of the Quaternary.
6.4 Medium sized carnivores
6.4.1 Red Fox (Vulpes vulpes)
European Red Fox, a small to medium-sized Canid, averaging a total body length of 1.75m and a height of 30cm, with an average weight of up to 3kg (Walker et al. 1975). The species is monogamous and breed during late winter. Breeding is often centred on a natural shelter, burrows or caves. Foxes are omnivorous and consume a wide range of foods but are known to prefer small mammals, birds, eggs, insects, and fruit and grass species. Identification of the species at Creswell relies wholly on morphological characteristics of their skeletal remains. Arctic Fox has also been reported by Mello (1875), and more recently the genera has been identified by Buckley et al. (2017) from DNA. Red Fox is known to be highly adaptive but in general terms prefers woodland or scrub habitats. In contrast, Arctic Fox is often an open ground preferring species and commonly specialises in predation of Grouse.
European Red Fox is known from 609 excavated bone fragments. Unfortunately, only 114 or can be relocated into a reconstruction of their distribution prior to excavation. Many of the specimens were originally stored in boxes containing the bones of smaller species and a number of these bones were unlabelled. The few bones that can be located within the deposits indicate that the distribution is extensive and seemingly concentrated within the upper levels. However, this is a tentative observation particularly as it is based on only 1% of the known remains. Despite the implication of their presence and the potential for disturbance in infilling sediments, they remain an enigma.
Red Fox population survival
Red Fox (Vulpes vulpes) remains are itemised in the digital archive and summarised for each stratigraphic level. European Red Fox are represented by bone fragments estimated to belong to 80 individuals.
The species remains are found throughout the sediment sequence but their overall survival is very low in all stratigraphic levels and averages around 4%. The explanation for this is not clear. For many levels, a small number of bones derived from a few individuals survive but in Level 6 a larger number of individuals, while represented by many bones, still have an individual very low survival rate. It is unlikely that this is an artefact of the excavation techniques as Armstrong routinely recovered very small micro-faunal remains.
The variation in surviving parts is illustrated in Figure 45. Destruction of the species remains is extensive. Bone frequency, compared to a live population equivalent, suggests a 90% to 95% destruction of bone. Despite being a small species known for its burrowing habits, it is probable that the distribution shown in Figure 45 is a reliable reflection of the temporal presence of Red Fox. The nature of the infilling sediments, which have an extensive limestone clast content, prohibits extensive disturbance as a result of burrowing.
Figure 45: Red Fox (Vulpes vulpes) temporal distribution and survival (source data: Digital archive)
Figure 46: Red Fox (Vulpes vulpes), body part selection (source data: Digital archive)
Variation in body parts is shown in Figure 46. Body parts are dominated by heads with very low numbers of limb parts and occasional occurrence of axial parts. This suggests that the accumulation is not due to carnivore activity. The variety of body parts, despite the low survival rate, includes all elements of the skeleton. This may indicate that the species was resident within the cave and the remains are the result of natural casualties.
Figure 47: Red Fox (Vulpes vulpes) attrition, bone damage (source data: Digital archive)
Attrition or damage to Red Fox remains is very distinctive. The results of analysis are illustrated in Figure 47. Complete bones number 47; there are no gnawed or carnivore damaged bone and over 420 bones are simply fractured. Causal factors accounting for the high frequency of fractured bone have not been subject to systematic study. It seems likely that the damage is the result of geomorphic events within the sediment body or from the trampling of exposed bone by the prolific faunal population known from the cave.
6.4.2 Arctic Fox (Vulpes lagopus)
This small carnivore is usually between 40-60cm in length with a body weight between 2.5-9kg. Modern studies indicate residence near or above the tree line and often in coastal locations. Diet is varied but concentrates on small mammals including lemmings, birds and carrion. Modern studies have shown that population size can fluctuate with that of Lemming species (Walker et al. 1975). Territories are known to be extensive and the species is known to wander over both arctic and woodland environments.
Magens Mello discovered a single complete vertebra within the red sand noted in his excavations (Mello 1876). The current whereabouts of the specimen is unknown. Little other stratigraphic information is available but it seems likely that this equates with the red sand sediment discovered by Armstrong and it would therefore relate to the stratigraphic levels 0 to 6. DNA fingerprinting has recently confirmed the existence of the species within the cave (Buckley et al. 2017).
Little direct stratigraphic and contextual evidence is available today but the species' ability to accumulate skeletal parts of their prey scattered around their residence areas has been recorded. It is unclear if this evidence occurred with the single individual recorded from the cave entrance.
This single species today enjoys an extensive holoarctic geographical range. The species averages 56-81cm in body length and has an average weight between 10-16kg (Walker et al. 1975). Individuals are largely nocturnal and are gregarious, often living in extensive burrow systems more or less continuously and which are extended as the group grows in size. Breeding takes place in late winter and birth, averaging 2 to 3 individuals, in early spring. A significant factor for this study is that modern burrows are known to be highly organised with sleeping areas, latrines, play areas and separate areas for the boars. There are numerous examples of burrows being cleaned on a regular basis. The species is omnivorous and feed on a variety of beetles, worms, slugs, voles and smaller prey such as young hares.
This species is known from 10 bones within the deposits and over half of these have sufficient dimensional details to incorporate in a reconstruction of their original pre-excavation position. The species has a random distribution of very low numbers of bone within the upper sequence. A single concentration is known from the area 64ft (19.2m) north of the entrance at a depth of 10ft (3m). The presence of Badger nearby in level 10 was used to suggest disturbance on vertebrate bones (Jacobi et al. 1998) and explain the earlier age. This is, of course, possible but a more likely explanation is that both bones were near the eastern wall of the cave where there was a known void (now filled with stalagmite) and where younger bones have fallen downslope toward the inner rear chamber (referred to by Jacobi et al. 1998 as the 'trefoil' chamber ). A second Badger bone, a first phalange, is known from the same area but from level 4ft (1.2m) depth and possibly 6ft (1.8m) above the dated bones. Armstrong records that this was the upper cavity of the chamber entrance. In other words, both bones (at least) did not have a coeval stratigraphic association with the remaining contents of stratigraphic level 10 depth. This example is a very good case of the dangers of using clearly labelled remains from excavations in complex environments and where association is assumed.
European Badger population (survival)
Survival of European Badger is shown in Figure 48. The occurrence of the species is unusual within this cave but it is known from bone fragments estimated to be derived from 15 individuals. Temporal distribution of this species is discrete and confined to a relatively large presence in stratigraphic levels 0 to 4, levels 7 and 8 and level 11.
Figure 48: European Badger (Meles meles) temporal distribution and survival (source data: Digital archive)
Remains contained within the lower stratigraphic levels have sufficient stratigraphic information recorded by Armstrong to indicate that they are from the rear of the cave passage adjacent to the 'inner chamber'.
European Badger attrition
Destruction of body parts (shown in Figure 49) is extensive with the relative exception of those from levels 2 and 3. Body part survival is dominated by limb fragments with some head parts. There are many similarities between this style of presence and preservation with that of Red Fox and with the comparative date known for European Badger at Steetley Cave. The multiple similarities suggest that this species was occasionally resident within the cave.
Figure 49: European Badger (Meles meles), body part selection (source data: Digital archive)
Analysis of body part variation and distribution suggest that there are two stratigraphically distinct clusters and these are illustrated in Figure 42. The youngest cluster in stratigraphic levels 0-4 has a diverse array of body parts, including axial parts of the skeleton. The style of bone preservation compares with that known from Steetley Cave which has been interpreted as a Badger den.
The skeletal body parts from 7-11 discovered near the entrance to the 'inner' chamber consist of a humerus, femur, radius and calcaneum, which may be from the same individual. The four bones have a vertical spread of 4ft (1.2m). This suggests that this one individual may have had a burrow or den in this area. Armstrong did not refer to the sediments in this area but there is photographic evidence of wall cavities within this area. There is also geomorphic evidence from later excavations that open cavities existed adjacent to the cave wall within the passage and chamber. In several areas, there is evidence of water flow and travertine formation down the eastern cave wall within cavities. In at least one instance, later travertine has formed upon the surface of a much older bone (a Mammoth tibia) within level 12.
Figure 50: European Badger (Meles meles), attrition - bone damage (source data: Digital archive)
Bone damage of the specimen's remains is minimal (Figure 50) and is dominated by complete (6 bones) and fractured bone (7) associated with an isolated occurrence of a gnawed fragment. The lack of serious fracturing and damage to these specimens supports the suggestion that this species is resident in layers 0-4.
6.4.4 Wolverine (Gulo gulo)
The single species of Wolverine (Gulo gulo) has a holoarctic distribution today and is known to prefer forest and tundra habitats. The species is a medium-sized carnivore of 60-80cm in length and usually weighs around 25kg with a dense body fur (Walker et al. 1975). They are a strong and agile carnivore that is able to drive much larger carnivores from their kills. They are solitary but have habitats that can range over an area of 300,000 hectares and which are shared by a male with a female. Diet is considerably varied but includes birds, and they are powerful enough to kill deer.
Three post-cranial bones discovered by Magens Mello during his 1876 excavations in the cave entrance represent Wolverine. There are no precise details of their stratigraphic context but they were discovered within the Red Sand, a deposit that equates with the stratigraphic levels 0-6 from the Armstrong excavation and are therefore likely to be of late Last Glacial age. The morphology of this species is distinctive and there is little doubt concerning the reliability of their identification, which seems to represent the remains of a single individual. There is little taphonomic information available but the species occurred in levels that included numerous fish remains.
6.5 Smaller carnivores
Small species of carnivore are known from 13 bones, which have been identified to the species Pine Martin (Martes martes), Polecat (Mustela putorius), Stoat (Mustela erminea) and Weasel (Mustela nivalis) (Figure 51). Their stratigraphic distribution is shown in Figure 51. Many of the identifications are on cranial bones, teeth or distinctive limb bones and there is little doubt that other specimens may exist that are as yet unidentified and which have been grouped within the collection of unknown rodentia. A further difficulty is that for some species, the small collection of bones appears to belong to the same individual, despite the fact that individual bones occur in different stratigraphic levels. This would seem to suggest that there has been considerable downward movement of their bones, which may be the result of burrowing activity or post-mortem movement of very small bones. Interpretation of their occurrence needs to be with some caution within each stratigraphic level. A full itemised inventory of small carnivore remains is available in the digital archive; the stratigraphic occurrence of species is described in spreadsheets for each level.
The species are aggressive and voracious predators that inhabit relatively small life territories and hunt a wide range of birds, rodents and amphibians. Without more precise details of their stratigraphic occurrence, an interpretation of their habits and ecology is difficult but their presence does indicate ground cover and they are associated with many avian and amphibian species that must include their potential prey.
Figure 51: Distribution of smaller carnivores (Mustelids) (source data: Digital archive)
Detailed discussion of the taphonomy for small predators is inhibited by the nature of the assemblage. The apparent dominance of cranial remains is without doubt an artefact of the analysis and identification difficulties of post-cranial remains of these species. There is no doubt that these were recovered during excavations and survive among the large collection of unidentified micro-vertebrate remains. The general complete state of micro-vertebrate remains, including small carnivores, suggests residence within or near the cave. Predation by raptors remains a possibility but there are no examples of the distinctive bite/killing damage to humerii or other post-cranial and basio-occipital parts.
6.6 Large Ungulates
6.6.1 Woolly Rhinoceros (Coelodonta antiquitatis)
This species belongs to a family with five living species inhabiting a wide range of varied habitats (Walker et al. 1975). It is largely assumed that the closest living relative is the African Black Rhinoceros (Diceros bicornis) and observations made in this study are in comparison to this species. This is a massive ungulate species, with head and body length known to be between 2-4.2m and with a shoulder height of between 1-2m. Adults can weight between 1000-3500kg. It is not known if the Quaternary species was solitary or lived in small groups; both arrangements are known within modern species. Breeding often produces a single young. Locomotion in Rhinoceros is poor, with speeds up to 45km per hour, but modern species have an acute sense of smell and hearing. Borsuk-Bialynicka (1973) has suggested that Woolly Rhinoceros was more ponderous than its modern relatives and may have been more susceptible to group attacks. Individuals are semi-nocturnal and show a marked preference for dense thickets of vegetation during daytime. They are attracted to shallow pools and rivers with sandy beds. The species is a grazer, feeding upon a wide range of grasses and succulents. Young animals are known to be targeted by larger carnivores while adults seem immune from most carnivore predation with the exception of humans.
The remains of Woolly Rhinoceros are not only well-preserved but are also well-recorded during the excavation. A total of 694 bones are known and 45% of these have sufficient dimensional information to be incorporated within the reconstructions. In addition, a further 239 bones can be allocated to levels 1-12 within the passage area. In effect this means that 79% of the specimens can be incorporated within the reconstruction. All specimens are illustrated by body part and these are colour-coded for crania, axial and limb parts. The species distribution is extensive and specimens occur in frequency from 33ft (10m) north from the entrance to 70ft (21m) north and within depths 5-12 ft (1.5-3.6m) below the stalagmite datum.
There are three separate concentrations, as follows:
Area 1: 62-70ft (18.6-21m) north from the entrance to depths between 3/5-10ft (0.9/1.5-3m); the distribution suggest that the sediments were inclined to the south. The levels contain 80 bones of this species and 66 are adult (28%) with 14 (2%) being juvenile. Only 4 bones show signs of gnawing or surface licking. Indicative of juvenile presence.
Area 2: 36-59ft (10.8-17.7m) north from the entrance and between depth 5-12ft (1.5-3.6m) below the stalagmite datum. This area had 61 bones, 90% of which were adult and 10% juvenile. The area also contained a bone with cut marks on one surface from the area 45ft (13.5m) north from the entrance at a depth of 12ft (3.6m). The 'passage area' had 127 bones of this species and 80% were adult and 11% juvenile.
Area 11: 35ft (10.6m) north from the entrance with depths 5-12ft (1.5-3.6m). This area has bones of this species randomly distributed within it.
The two main concentrations are within the passage and rear of the cave and the areas coincide with the distribution of both Neanderthal assemblages and the distribution in the rear of the cave. Area 2, with a single cut bone, clearly demonstrates Neanderthal presence within the passage.
Woolly Rhinoceros population survival
This large species is known from a total of 694 bone fragments estimated to belong to 40 individuals, which are shown in Figure 52. It is very likely that further vertebrate bones are derived from this species but the reduction by extreme fragmentation presents difficulties in identification to generic or species level. It is likely that further fragments may be identified among the large collection of bone flake material. The remains are relatively prolific and initially appear within stratigraphic levels 11 and 12. Their frequency steadily increases to a peak in levels 6 to 9 and is followed by a steady decline in numbers. The temporal occurrence almost exactly replicates that of Hyaena distribution. Despite the large size of the species, bone survival is low in comparison with an original live population. The remains include a number of very old individuals associated with a larger frequency of sub-adults, which may indicate predation by hunting and scavenging of carcasses.
Figure 52: Woolly Rhinoceros (Coleondonta antiquitatis)- temporal distribution and survival (source data: Digital archive)
Individual survival of Woolly Rhinoceros is shown in Figure 52. They are present in all levels but most of the fossil population occurs within levels 8-14, with a particular peak in stratigraphic level 10 where relatively large numbers of skeletal parts survive.
Woolly Rhinoceros attrition
Figure 53: Woolly Rhinoceros (Coleondonta antiquitatis), body part selection (source data: Digital archive)
Variation in the survival of skeletal parts contrast quite markedly with the pattern of overall survival (Figure 53). The remains are dominated by head and limb parts with very low numbers of axial parts. The pattern of survival clearly confirms that the kill or death site of such a large species is elsewhere and that limb and skull parts have been transported into the cave. The presence, in low numbers, of axial parts suggests that there has been little competition at the 'kill' site. The significant peak in stratigraphic level 11 is directly associated with both Neanderthal and Hyaena use of the locality. Both cut (by Neanderthals) and gnawed bones (by Hyaena) occur in the level. The increase in the survival of head parts in stratigraphic levels 9-6 coincides with a decline in Hyaena numbers and an increase and diversification in Middle Palaeolithic artefacts. This is a significant association which is discussed below in considering Neanderthal food procurement.
Figure 54: Woolly Rhinoceros (Coleondonta antiquitatis) - a distal humeral fragment showing Hyaena fragmentation, gnawing and salival surface polish, Pin Hole Cave (Image courtesy of Manchester Museum)
Bone damage and attrition (Figure 55) is very distinct for this species. There is massive destruction of bone remains. The persistence of gnawed and fractured bone in stratigraphic levels 7 and 12 is a direct indication of carnivore activity in the form of bite marks, chewing marks and surface licking/polishing of bone. The humeral shaft shown in Figure 54 is typical of several known from the cave. Distinctive Hyaena gnawing has removed both epiphyses and the edges bear traces of bite marks by the same species. The spongey bone interior, typical of the species' limb bones, has also been removed. The whole bone surface is polished as a result of licking, probably by juvenile Hyaena. Such pieces are numerous within the cave and have, in past studies, been mistaken for artefacts (Kitching 1963). The specimen also indicates the presence of adult Hyaena, whose developed dentition is able to transport and fragment such a robust bone.
Figure 55: Woolly Rhinoceros (Coleondonta antiquitatis), attrition, bone damage (source data: Digital archive)
The occurrence of bone fragmentation of this species in overall terms also replicates Hyaena temporal distribution and is illustrated (Figure 55). There is, however, a major break within this pattern, which is clearly seen in levels 8 and 12. These levels contain numbers of complete and undamaged bones, one of which (in level 12) bears surface cut marks. These levels contain extensive archaeological evidence of the Middle Palaeolithic. It is difficult to explain the undamaged nature of the pieces, which are present within such a destructive carnivore environment. There is no evidence of rapid complete burial to explain their condition.
Stratigraphic level 12 also contained a scapula fragment of this species, which has a well-preserved coracoid process and associated rib but which has been heavily damaged along the blade. The fragment has edge damage near the coracoid which consists of a tooth/bite mark of Hyaena surrounded by a surface of incised tooth or chewing marks. The lateral surface of the scapula has a series of individual surface scratch marks along its edge, again the result of Hyaena. The presence of these pieces and their condition, one of which bears direct evidence of human agency, are likely to be an indication of Middle Palaeolithic use of the species as a resource and at a time when Hyaena was seasonally absent.
Figure 56: Woolly Rhinoceros (Coleondonta antiquitatis) mandible from stratigraphic level 9 (Image courtesy of Manchester Museum)
A large fragment of Woolly Rhinoceros mandible (Figure 56) was discovered within stratigraphic level 9 during Armstrong's excavation. The piece contains a well-preserved dentition, indicating it belonged to a juvenile. The preserved mandible represents approximately 70% of the whole jaw. The obvious fracture is vertical and appears in part to be not recent. The fracture itself has sharp 'green stick' edges. There is no evidence of percussion on the piece. The right lateral surface is preserved and the terminus has distinct carnivore gnaw marks along its edge. It is possible that similar damage was present on the left lateral edge but more recent breakage has destroyed much of the evidence. The complete piece is well within the carrying capacity of several carnivores. The absence of gnawing damage or bite marks on the remainder of the bone surface, particularly the medial edge of the mandible, within the context of the cave makes the piece unusual as it has been transported some 63ft (19.2m) into the rear of the cave. Within this general area, but at a lower level (stratigraphic levels 11 and 12) there are examples of human-cut Woolly rhinoceros and Mammoth bone that have no gnawing damage. All of the levels are within the range of deposits that contained lithics from the earliest Middle Palaeolithic. The evidence demonstrates that, in general terms, the presence of Hyaena and human activity within the cave is not mutually exclusive at this point in time but may represent differing seasonal activity.
Woolly Rhinoceros, cut or scratch marked mandible
Figure 57: Woolly Rhinoceros (Coleondonta antiquitatis) mandible fragment with surface marks. (Drawing by Andy Palmer; see also digital archive)
Stratigraphic level 12 yielded a large mandible fragment from Woolly Rhinoceros and is illustrated in Figure 57. This specimen is a fairly well-preserved mandible and dentition but is fractured behind the premolar on the left side. The right medial end of the mandible has a clean break that may have signs of carnivore teeth marks. The surface of the right pre-mandible has at least four near-vertical marks on the surface that are clustered together in the lower area of the mandible. The identification of causal factors is made more difficult by the observation that this location on the mandible is exactly where large carnivores such as Hyaena attempt to fragment mandibles. There are no other carnivore teeth, gnawing or salival polish marks on the mandible surface. The impressions are tightly clustered within one area and have fairly sharp edges, although they are of greater width than the more usual incised marks known from the cave. They are, therefore, interpreted as cut marks and the result of Middle Palaeolithic or Neanderthal group activity and perhaps the result of attempts to remove the tongue.
Cut-marked Woolly Rhinoceros tibia
There is also unequivocal evidence of human presence from level 12 in the form of a clearly cut-marked Woolly Rhinoceros tibia, shown in Figure 58. The item is clearly marked 12/P in Armstrong's handwriting, which indicates that it is derived from a depth of 12ft (3.2m) within the passage area. The size and lack of a fused distal epiphysis indicates that the bone is derived from a juvenile individual. The distal epiphysis is missing and the unfused end is corroded or eroded, which may indicate that the better-preserved proximal end was buried within the contemporary sediment. The lateral mid-shaft area of the bone has at least one group of clear and oblique cut marks. The seven short incisions are mid-shaft on the lateral surface and just below the anterior border (illustrated in detail in Figure 59). The occurrence of the incisions as a closely packed group in one location may indicate a single attempt to sever the prenoeus muscle, which originally would have occupied the whole lateral length of the tibia. Such a grouping may also indicate that this is a specific activity and not one of generally removing muscle. Given the sheer size of the bone, its likely partial burial, the associated presence of thirteen struck quartzite flakes and a biface of Middle Palaeolithic type, it does suggest that it has not moved fa. As such the piece seems to be a very good candidate for age estimations of the earliest Middle Palaeolithic within the cave. The lack of carnivore damage may also indicate an absence of such a population.
Figure 58: Cut-marked bone from stratigraphic level 12, general view (Jenkinson 1978b (Image courtesy of Manchester Museum)
Figure 59: Detail of cut marks on the mid lateral border (Jenkinson 1978b; Image courtesy of Manchester Museum; also see Digital archive)
6.6.2 Hippopotamus (Hippopotamus amphibius)
This species is well known from locations within Eastern England where it is often found with fluvial deposits. It is also commonly associated with the last interglacial. The bone collection in the University of Oxford Museum contains a single left molar, consisting of fractured pieces that have been reassembled and stuck together. It is associated with a museum catalogue number and archives that indicated it was donated by Leslie Armstrong and is from his Pin Hole excavations. In addition to an attached label, 'Pin Hole Cave, ultra-violet light reveals a clear mark 'B' upon the tooth surface'. This may indicate a location 'B' on an accompanying stylised section of the cave sediments drawn by Armstrong. The tooth has not previously been described in studies of the cave and has never been referred to by Armstrong. In the same collection of items, a Mammoth (Mammuthus primigenius) deciduous molar is labelled 'A', which appears to refer to a location in the same drawn section of Armstrong. In a later study, Jacobi et al. (1998, 35) have stated that the tooth is from Africa and not from the cave. Unfortunately, this is not supported by any published evidence despite its importance for understanding the age of deposits in the lower area of the cave.
The presence of this single tooth suggests that it may be derived from inter-glacial sediments within the cave that pre-date the current suggested age estimations. There is no other available evidence from the cave that would clarify the context of this find. The many thousands of associated palaeontological remains are all of species generally accepted as last glacial age. Much older sediment areas are known from areas of the roof deposits directly above this location. Uranium series age estimation for these sediments suggests an age of last interglacial age (Rowe et al. 1989, UEA101). Despite the age, it is very unlikely that the tooth is derived from these older and higher sediments, particularly as it is difficult see how a fossil of this type could be deposited in such a location originally. I am inclined to give Leslie Armstrong the benefit of the doubt, particularly as his many thousands of other observations within this site have proved to be reliable, once his methods are understood. I suggest that the tooth is a genuine find and was collected (from one of the many deposits known in Eastern England) by Neanderthals, as a raw material or curiosity and has been incorporated in sediments in the cave rear that also contain many other elements suggesting their presence.
6.6.3 Woolly Mammoth (Mammuthus primigenius)
The species is extinct and comments are based upon studies of its modern living relative, the African Elephant (Kühme 1963; Stuart 1982), particularly from Tanzania. This is a large species and can reach 4500kg in weight (Laws et al. 1975). Groups are fairly small and family based. Larger groups tend to be an agglomeration of families. They are matriarchal and the group's leader is usually the dominant female. Adult individuals are largely immune from predation but large carnivores are know to predate on juveniles. Food consumption is significant and can be up to 4% of body weight, daily. They are known to migrate regionally and to visit regularly exploited sites including caves. Sutcliffe (1985) cites an example where modern elephants entered a cave to lick salt from the walls and in doing so over the years considerable altered the area of cave itself. Frozen remains of Mammoth have demonstrated that the species had a thick pelt and layers of subcutaneous fat. Although discoveries of their remains are plentiful, little is known about their group size. African elephants can travel at between 20-30 miles per hour. Adult individuals drink up to 40 gallons of water per day and large amounts of time are spent in the vicinity of rivers, waterholes and other water resources.
They display considerable group defensive behaviour when confronted by predators. Modern field observations from Southern Africa have documented at least one incidence of a family group of Elephant that were attacked by four armed hunters. In this instance, the family group attacked and killed the assailants during which the lead male was shot 34 times. Natural and accidental deaths in modern populations can often produce a landscape that is randomly covered with the species' carcasses and they are an important scavenging resource. Fossil localities are often associated with fluvial and marshy environments. When such localities contain associated lithic remains, it is often assumed that they represent kill sites.
Figure 60: Woolly Mammoth (Mammuthus primigenius) distribution and survival (source data: Digital archive)
Woolly Mammoth is known from the presence of 227 bone fragments, estimated to be derived from 30 individuals. Figure 60 shows that they occur throughout the cave's history, in 12 of the 30 stratigraphic levels. There are two significant increases in bone frequency in stratigraphic levels 11 and 6. For the suggested 30 individuals represented within the cave, the level of destruction of skeletal remains is enormous, with only 1-2% represented by remains within the cave. Such a low rate of survival probably reflects the fact that many of the skeletal parts were never transported into the cave. Selection of bone, for whatever reason, was achieved outside of the cave environment.
Woolly Mammoth attrition
Figure 61: Woolly Mammoth (Mammuthus primigenius), body part selection (source data: Digital archive)
The frequency of body parts and their survival is illustrated in Figure 61. Analysis of the source data shows that many of the fragments are derived from cranial parts and many of these are remnants of teeth that have often been shattered into numerous pieces. Under these circumstances it is difficult to establish association between the respective parts of the same tooth, which itself makes estimation of the minimum number of individuals precarious. In any event, this over-estimates the frequency of teeth fragments and survival.
Distribution of body parts is within practically all levels and includes multiple tooth fragments in association with limb parts. Stratigraphic levels 1-7 also have a high number of dental parts with isolated limb fragments. Many dental parts are tusk or ivory pieces, which occur in levels with human occupation. It has been assumed that Woolly Mammoth were extinct within Britain during the period that these sedimentary levels accumulated. Clearly all body parts have been transported into different parts of the cave. Figure 63, a juvenile, known from the 1874 exploration, was discovered in the cave entrance (Mello 1875) and may have been carried into this part of the cave. Much of the material from within the cave is that of juvenile individuals.
Figure 62: Woolly Mammoth (Mammuthus primigenius), attrition, bone damage (source data: Digital archive)
Within the massively reduced and destroyed skeletal parts, molar teeth (predominantly adult) are significantly more frequent and many of these have been fractured (Figure 62). Survival may indicate a deliberate collection of these parts, which have poor dietary value from a carnivore's viewpoint but which may have provided a resource of unknown use for human groups. Circumstantial evidence for this view is that such parts are more numerous in stratigraphic levels 6 to 12, which have Middle Palaeolithic archaeological evidence. Level 12 contained a single, undamaged and ungnawed, juvenile Mammoth tibia that has surface cut-marks. Similarly, such teeth have an isolated distribution in levels 2 and 3 characterised by Upper Palaeolithic evidence.
A partial record of finds was written by Leslie Armstrong within his excavation notes. He records a number of interesting occurrences of this species, three of which are significant.
At 5ft 6in. depth (1.56m) (in the passage?) a Mammoth tusk fragment was recovered in association with a unifacial quartzite tool
At the same depth, a small worked ivory fragment was recovered in a 'pocket' of the eastern wall. This may be one of the pieces illustrated in the archive (A9, A10, A11, A12)
At a 5ft depth (1.5m) a Mammoth tooth was recovered in direct association with a Hyaena mandible fragment and a quartzite artefact
The stratigraphic details are not precise but the information available indicates that at least three worked ivory pieces occur within an area containing quartzite and flint lithics of Middle Palaeolithic type. The occurrence of at least one piece in a wall cavity is an interesting detail and points to a deliberate attempt to store it. Various depths have been recorded for these finds which suggests that they may be of a younger age. The finds come from a region where the passage and chamber deposits can be shown to slope eastwards, making correlation with the western area deposits difficult. At least one older tibia located within this area has a stalagmite coating, which seems to be a post-depositional deposit and has been dated to the Late Glacial.
There is further evidence of the possibility of Mammoth ivory working by Neanderthal groups from the deposits 5ft 6in (1.56m) below the stalagmite floor. These are recorded by Leslie Armstrong in his field notes. He recorded a quartzite tool associated with four shaped ivory pieces that were within and associated with an alcove on the eastern cave wall.
Figure 63: Juvenile Woolly Mammoth (Mammuthus primigenius) mandible which was discovered within stratigraphic level 9 within the passage area of Pin Hole Cave (Jenkinson 1978b (Image courtesy of Manchester Museum)
A single specimen of a juvenile Mammoth mandible was discovered within stratigraphic level 9 (Figure 63), which also contains Middle Palaeolithic lithics. The piece has two clear bite marks on the medial surface and an adjacent area of surface scratching, which indicates the presence of juvenile Hyaena.
Cut-marked Mammoth bone
A large and near complete tibia and was discovered by Armstrong in Layer 11. Its morphology suggests that it is a right tibia of Mammoth (Mammuthus primigenius) (Figure 64). The proximal epiphysis is absent and may have been unfused, suggesting that it is from a juvenile. Bone flakes are exfoliating from the shaft surface and a longitudinal split in the bone is present. It is not clear if these features are contemporary with deposition or are later damage from storage. Seven clear incisions are present mid-shaft and extend along the anterior medial surface (see Figures 65 and 66). Their location is in the region that was originally covered by the peroneus muscle and may reflect attempts to sever this. In common with the previously described example, the grouping and coincidence of location may indicate a specific single activity related to severing a single muscle and not one of general defleshing.
Figure 64: Drawing of Woolly Mammoth (Mammuthus primigenius) tibia showing surface incisions on the central shaft (Drawing by Andy Palmer)
Figure 65: General view of the cut-marks on a Woolly Mammoth (Mammuthus primigenius) tibia from stratigraphic level 11 (Jenkinson 1978b, Image courtesy of Manchester Museum)
Figure 66: Detail of cut-marks on the mid-anterior surface (Jenkinson 1978b, Image courtesy of Manchester Museum)
Predation of the species appears to be confined to juveniles but the identity of the predator is difficult to identify. Some specimens are gnawed and have bite marks from Hyaena and at least two pieces have cut-marks with no evidence of carnivore damage. Both forms of evidence are also found within the same stratigraphic layer, which indicate predation by both humans and Hyaena. Hyaena have not damaged the tibia. This is a clear indication of Neanderthal presence within level 11.
6.6.4 Wild Pig (Sus scrofa)
This is a nocturnal species that shows considerable size variation in modern populations, generally becoming larger in cooler, temperate environments (van Den Brink 1977). Individuals live a solitary existence except in the breeding season when they congregate in small groups. The species shows a preference for mixed woodland with daily access to water, particularly in marshes. Residence is in shallow burrows or rocky ground and modern populations are not normally found in caves.
Wild Pig is relatively rare within the deposits and only 12 bone fragments are known, estimated to be derived from 4 individuals. The stratigraphic distribution is described in the digital archive. The most significant stratigraphic distribution is from stratigraphic levels 11-12. Analysis of body parts indicate that the remains are predominantly cranial with a few vertebrae fragments. Bone damage or attrition indicates that all are fractured, with no evidence of gnawing. The overall number of skeletal parts is very low but their occurrence in stratigraphic levels 11-12 and 6 are almost certainly the result of Middle Palaeolithic activity. Their occurrence in stratigraphic levels 1-2 may be the result of Upper Palaeolithic activity.
6.6.5 Wild Horse (Equus sp)
Ethnology of Horse (Equus sp)
The identification of this species remains ambiguous. Fossilised remains, particularly from Creswell, vary considerably. Many studies have suggested that the Quaternary species is closely related to the modern day Prezewalskii horse (Equus prezewalskii) on the basis of its stocky form and distinct mane, both known from fossilised remains and Palaeolithic artistic depictions. A more robust species, the European tarpan (Equus caballus) also seems to be present within the fossil record and was alive within the historic period. It is possible that separation upon the basis of morphological variation within skeletal remains is not sufficiently diagnostic and that a more clear definition may be available from DNA studies.
Whatever the view of species, it is clear that these animals live in large groups, are grassland grazers and prefer open plains. Head to body length for the species varies from 1.8-2m and height to the shoulder is 1-1.2m (Walker et al. 1975). Body weight can vary but averages 350kg. The species is extremely mobile and wary of predation attempts. It is not known if this was an aspect of Quaternary Horse behaviour or if it is an acquired behaviour resulting from extensive human predation within historic times. Adult animals are largely immune from predation by large predators. Horse populations are known from modern studies to occupy large home territories and it is essential that they have daily access to water. Horse remains from Pin Hole are itemised and are summarised for each stratigraphic level in the digital archive.
The known collection numbers 463 bones and 346 of these have sufficient dimension details to be incorporated within the reconstruction, estimated to be derived from 48 individuals. The main concentration of Wild Horse remains occurs within 63-66ft (18.9-19.8m) and within 5-10ft (1.5-3m). A total of 41% of the known remains occur within this area. Specimens that can be traced to the 'passage' levels 9-12ft (2.7-4.3m) add another 25% to the total reconstructed distribution. Wild Horse remains have been grouped by age. Within an area slightly larger than the main concentration, adults are 47% of the population and associated juveniles are 36%. Remains that can only be sourced to the passage within this comprise 39% adults and 60% juveniles. The occurrence within this area of the cave is very distinctive in that a significant number of the population are adult females with associated foetal and juvenile remains. The concentration coincides with that of the Later Middle Palaeolithic lithic assemblage.
Wild Horse population survival
The remains of this species are present in virtually all levels of the cave (Figure 67). There are notable peaks in its distribution in levels 11-12 and levels 5-9, which coincides with Middle Palaeolithic lithic remains. The peak occurrences and interrupted distribution also contrast with the temporal distribution of the major carnivores. Studies by Turner (1981) have suggested that the Pin Hole population is dominated by older females, which in their natural habitat would have been more prone to predation by large carnivores.
Figure 67: Wild Horse (Equus sp) temporal distribution and survival (source data: Digital archive)
In common with other species known from the cave, the surviving remains represent a small part of those expected within a live population and indicate that bone selection and destruction averages over 90%. Individuals occur in low numbers, averaging 2 to 3 animals in many stratigraphic levels with the single exception of level 12 where up to 10 individuals may be present. Most of the bone is derived from adult or sub adult individuals. Three stratigraphic layers (6 and 11-12) have relatively higher levels of bone survival than the other layers). These levels also have evidence of Middle Palaeolithic use.
Wild Horse body parts
Wild Horse skeletal parts are summarised in the digital archive and illustrated in Figure 68. Cranial parts total 277, with only 174 limb fragments and 12 axial parts. Such distinct selection (and survival) of these parts clearly demonstrates that not only were most individuals prey but also that the body parts found (approximately 30% of the live total) were transported into the cave. Many of these bones have survived with virtually no associated axial parts in levels 2-12 with particular frequency in levels 5-7 and 9-12. These levels all have Neanderthal artefactual evidence. The lower layers also have large number of Hyaena known to be in sporadic residence.
Figure 68: Horse (Equus sp) body part attrition (source data: Digital archive)
Figure 69: Wild Horse (Equus sp), bone damage
Over 80% of bone is fractured, associated with a large number of complete undamaged bone (419 bones). There are very low frequencies of gnawed or fractured and gnawed bone (44 bones). The general pattern of bone damage is quite different from that of other species such as Woolly Rhinoceros and Mammoth, where gnawed bone occurs in relatively significant numbers. In this context gnawed bones resulting from Hyaena predation are rather rare. One of the few examples, a maxillary, was excavated within stratigraphic level 6 and this is illustrated in the digital archive. The fragment consists of a pre-maxillary with well-preserved incisors, which has been fractured from the main skull just below the nasoid. The bone edge has two very distinct carnivore molar impressions penetrating the fragment.
It seems likely that this style of bone damage (and preservation) may be the result of human butchery techniques, which result in large numbers of fractured but ungnawed bone. This suggests that stratigraphic levels 11-12 and 5-6 are important temporal occurrences of human use of the cave. The survival and variation in Wild Horse remains suggest that these were of considerable interest to human groups and were of especial importance in levels with Neanderthal use in stratigraphic layers 6-12. While these layers seemingly contain evidence of Hyaena, there is sporadic but infrequent evidence they preyed on Horse.
6.6.6 European Bison (Bison sp)
Identification of Bovid species present during the Quaternary is notoriously difficult, particularly for post-cranial remains, and the situation is made more difficult by the presence of sexual dimorphism in most species (Stuart 1982; Walker et al. 1975). There appear to be two genera present, Bos and Bison. Many studies suggest that the Quaternary European species is likely to have been European Wisent (Bison bonasus), which is known from deposits of historic age and documents the slow extermination of the species to a current small and protected population. European Wisent is thought to prefer woodland habitats rather than the open plains favoured by other Bovid species.
Body length is usually between 2-3.5m, with a shoulder height of 2.5-2.8m and a body weight that can vary between 450kg to 1350kg (Walker et al. 1975). Average herd size for the European Wisent is not known but may compare with the enormous herds documented for the American Bison (Bison bison), which at times have numbered hundreds of thousands. For modern populations, the breeding season is between July to September with a gestation period of nine months. Although the species is fairly agile and mobile, it was probably one of the slower ungulates present within the Quaternary. Modern populations are known to undertake lengthy migrations.
European Bison remains consist of 289 specimens known from Pin Hole Cave. A total of 127 of these are used in this reconstruction, representing 54% of the known remains. The spatial distribution of European Bison remains occur in low numbers, spread evenly between depth levels 3-12ft (1.5m-36m) for most of the cave's length. There are two distinct concentrations of distribution as follows:
Area 1: within 63-72ft (18.9-21.6m) north from the entrance within depths 3-6ft (1.5-1.8m) below datum. This area contains 36 bones, all probably adult. Thirty specimens (a further 10% of the total remains) are known from the passage, which represent basal levels 11 and 12 in this area. The occurrence suggests that the bones are within the sediment mound in the rear of the cave and may be inclined down the southern facing slope. The location of this concentration is very similar to that of other large ungulates from the cave rear.
Area 2: within 31-56ft (9.3-16.8m) north from the cave entrance and within depths 3-6ft (1.5m-1.8m) below datum. This area contains 54 adult bones, which represent 18% of the reconstruction population. The distribution and body part type is unusual in that cranial parts are almost absent. Bones also occur at the interface of Middle and Upper Palaeolithic assemblages in the front area of the cave.
European Bison population survival
The bone fragments are estimated to belong to 19 individuals. It is certain that this analysis has under-represented the species presence. This is due to the presence of large numbers of fragmented vertebrate bones, which lack sufficient morphological features allowing firm identification to this animal but which nevertheless may derive from it. Many of the remains are also from post-cranial parts of the skeleton, which are extremely difficult to allocate to a particular species on morphological criteria alone. All of the examined remains are therefore grouped within the Bovid genera. It has not been possible to distinguish conclusively any of the remains to species level. In effect, the combination of species size variation and sexual dimorphism, largely of post-cranial bone, means that they could be derived from one or two different species belonging to the Bison genera with includes both Bison priscus and Bison bonasus, the European Wisent.
The remains of these genera have a very distinct temporal distribution, shown in Figure 70. Its first appearance is of several individuals in stratigraphic layer 16, and these may in fact be a single individual. Within the upper levels of the cave, there are discrete distributions in levels11 to 13 (Middle Palaeolithic), levels 7 to 9 (Middle Palaeolithic), levels 4 to 6 (Middle and Upper Palaeolithic) and levels 1 to 3, (Upper Palaeolithic). It is quite clear that the distribution coincides, stratigraphically, with human occupation evidence and contrasts with that of the major carnivores already discussed.
Figure 70: European Bison (Bison sp) temporal distribution and survival (source data: Digital archive)
Bison remains from Pin Hole are itemised and summarised for each stratigraphic level (Digital archive). Estimations of the minimum number of individuals, shown in Figure 70, suggest that the species is consistently present throughout the entire sequence and in relative terms the level of survival of their remains is of a very low frequency. The extensive destruction of remains from a specific selection process or on-site attrition is, however, fairly uniform within the sequence. Stratigraphic level 2 has a 10% survival rate, which, while low, is twice the rate of anything else within the sequence. Similarly, minor increases are known within levels 5-6.
Bison attrition
The distribution and variation of body parts for this species is shown in Figure 71, where it is immediately obvious that the preservation contrasts with practically all other species known from the cave. The size of the species concerned rules out any possibility of a natural passage into the cave and the variation in body parts therefore clearly indicates a selection process where both limbs and axial parts are selected. Cranial and head parts are present but occur in very low frequency. The possibility of selection by large carnivores can be discounted; despite their presence, with the exception of stratigraphic level 5, significant peaks in frequency coincide with the occurrence of archaeological evidence for both the Middle and Upper Palaeolithic.
Preservation of body parts is distinct and different from many other species within the cave (Figure 71). Unusually, there are very few head parts and the few that do occur are mostly teeth, with horn parts and other cranial bone being very rare. Limb and axial parts form the most frequent remains. The relatively high survival of individual European Bison remains and the distinct pattern of selection clearly suggest strong control over a kill site, indicating that the more nutritious parts of the body were transported into the cave. The body part remains indicate that carcasses were butchered or separated and limb and axial areas were brought into the cave. This suggests a pattern of hunting rather than scavenging. The contrast with evidence from levels known to have Hyaena residence suggest that human groups were responsible for this accumulation. Stratigraphic levels 9 and 11 have evidence of Neanderthal occupation and Hyaena are also present within these levels.
Figure 71: European Bison (Bison sp), body part selection (source data: Digital archive)
The upper stratigraphic levels 1-5 have extensive archaeological evidence from the Upper Palaeolithic and this is associated with evidence of a similar selection of body parts (limbs and axial remains). The carnivore Hyaena is absent from these levels and there is evidence of a small European Wolf presence.
Attrition - bone damage
Attrition of Bison remains is described in terms of bone damage type within Figure 72. The results of this analysis are distinct and unlike similar analyses for other species. A significant amount of the material is fractured (93 bones) and a large amount is gnawed (59 bones). The presence of complete and undamaged bone is confined to half of the stratigraphic levels. While the general mode of occurrence is similar within the sequence, there are large differences in the frequency of damage type between each occurrence and this is particularly noticeable for the presence of fractured and gnawed bone. The occurrence of complete bone appears to increase through time and is at its most abundant in the upper stratigraphic levels of the sequence. Within each distribution cluster, the occurrence of gnawed material appears in isolated peaks of frequency, indicating a relatively rapid change in circumstance over preceding levels.
Figure 72: European Bison (Bison sp) attrition, bone damage (source data: Digital archive)
Such peaks may indicate carnivore appearance following their relative absence in underlying levels. Description of Hyaena presence and residence within the earlier levels of the cave have drawn attention to their initial abundant occurrence and slow decline in frequency. Mid-sequence is associated with a description for European Wolf, where there is a dramatic increase in frequency within the upper parts of the sequence. Both species have the ability to fracture bone. In the case of Hyaena, it is clear that this is possible with bone belonging to some of the largest species, such as Woolly Rhinoceros, which are known from the cave. Despite the difficulty of reliably identifying their respective gnawing activity, it does seem that the reduction of vertebrate bone by Wolf is not as developed as that known for Hyaena. It may therefore be the case that one species' activity has simply been replaced by another within the sequence and that such activity by both species has an interrupted occurrence that is also associated with unequivocal evidence of human presence in the form of lithic assemblages. A single Bovid radius from stratigraphic level 6 has surface cut-marks indicating direct human interest. The specimen was originally excavated within the area of 53ft north from the entrance and is therefore within the area of Middle Palaeolithic artefacts. The depth of discovery suggests that it is derived from the Later Middle Palaeolithic.
Within stratigraphic levels 10 to 12, gnawed Bison bone is present within level 12 and is absent from the overlying two levels (0.6m of deposit) and where evidence of Middle Palaeolithic lithics is present. This contrast may indicate evidence of the cessation of Hyaena residence and the beginning of human use. A similar phenomenon is present within stratigraphic levels 6 to 9, but in this instance there is variation level by level within the occurrence. These levels also have evidence of a later Middle Palaeolithic use but which is less clearly defined. Nevertheless, the distribution of gnawed bone is clumped, with an absence within intervening levels and as such may indicate the same process but of more complex form. Similar observations can be made for stratigraphic levels 1 to 4 where there are two separated distributions within levels containing Upper Palaeolithic lithics. Although the evidence is circumstantial, it does suggest that there are at least two separate occasions when human groups were present.
Within stratigraphic levels 1 to 9, analysis of species distribution indicates that Hyaena frequency declines significantly and is accompanied by a dramatic increase in the present of European Wolf (see Digital archive). It seems likely that Hyaena and human groups made use of the cave at different times. There is direct evidence of human interest in Bison in the form of a photograph taken at an unknown location within the passage. The photograph shows an in situ Bison humeral shaft with a blade tool resting between the distal epiphysis. It is unfortunately not possible to identify this specific bone within extant collections but the presence of the blade indicates that it is derived from the upper stratigraphic levels within the passage.
6.6.7 Reindeer (Rangifer tarandus)
The single species of Reindeer (Rangifer tarandus) is native to the arctic region. The species displays sexual dimorphism, with body length varying between 1.2 to 2.2m, shoulder height between 2.2 to 1.4m and body weight of up to 318kg (Walker et al. 1975). Both sexes have antlers, which can vary greatly in form, and they have specific adaptions for arctic environmental habitats. Herds can be very large.
Ethnology of Reindeer (Rangifer tarandus)
This species is consistently present on European Quaternary sites and, allowing for variation in density in any one-place, they are often represented by large numbers. In many cases, the species occurs with direct evidence of palaeolithic use. While most archaeological evidence is in the form of stone tools, there is much evidence of bone and antler tools, engravings and paintings. The species is also commonly found in association with Horse.
Modern populations have been extensively studied (Banfield 1954), particularly the nature of their relationship with human groups. Many studies have demonstrated their relationship with palaeolithic groups either on individual sites or regionally, and the ecology of modern Reindeer has often been used as a parallel for palaeolithic practice and procurement systems. This approach suffers from both the significant palaeogeographic changes that have taken place during the Quaternary and that few modern populations are unaffected by human control, often upon a very local basis.
This reassessment makes use of the research upon the barren-ground herds of Northern Canada (Banfield 1951; Cosewic 2016; Gordon 2004; Frame et al. 2008), which have limited contact with human groups and range over a vast area of around 9000km² in any given annual cycle. The details of Reindeer ethology are considered in some detail, particularly as their specific behaviours may offer an understanding of their Quaternary circumstances and palaeolithic procurement systems at Pin Hole Cave.
Following their widespread occurrence during the Quaternary, the species has declined and is now restricted to their preferred biome in Holarctic regions. Their distribution and occurrence within these regions is accompanied by considerable variation in their size, behaviour and biome preference. In modern populations, individual animals vary from 1.2 to 2.2m in height and can weigh between 200 to 718kg. All individuals have a seasonally varying coat as protection against cold and wind (Walker et al. 1975). Both sexes grow antlers, which are shed by males in autumn and females in late winter or spring. The antlers are sufficiently morphologically different to allow identification of each sex. Current populations, such as the barren-ground herds, are known to achieve a herd size of approximately 250,000 individuals. Much of the actual travel is undertaken during the summer as one herd spreads over a large area. Individual groupings take place during the breeding season and in spring, during the calving season; females will often form smaller groups with the males in separate groups ranging over a wider area.
Many known modern groups display considerable agility and move into new areas, often developing variation in types. The success of such movements is heavily affected by interspecies competition which, when it is developed, results in a clumped or interrupted distribution. Movement is often along watercourses and river valleys and there seems to be a dislike of high ground. Migration of this type is annual and aimed at locating fresh grazing and the avoidance of parasites and predators. Migration is often swift, takes place during the summer months with a view to locating higher cooler ground, which offers a more tranquil environment for calving. Summer grazing is often generalised but concentrates on birch and willow scrub. Winter grazing is located on higher more exposed ground where a preference is shown for mosses and lichen, particularly reindeer moss. Many of these plant species have low density and are slow growing, giving rise to the need to forage over large areas and for substantial breaks in yearly grazing to allow the vegetation to recover.
Summer movement is strongly affected by parasites and predators. The existence of parasites represents one of the principal threats to herd safety. Herds suffer from parasitism from Warble Fly (Hyenomipia trompe) and Bot Fly (Gasterophilus intestines), which are often concentrated in warm and wet river courses. Both species attack adults but will also particularly target juveniles. Warble flies utilise the exhaled carbon dioxide to locate and attack, and they lay eggs in the lips and nasoid region where the developed maggots give rise to cardiovascular problems. In contrast, bot flies locate individuals by urine detection and colonise the rectum and feet glands, resulting in inhibited movement in the host. Both infestations have serious implications for an animal that relies upon rapid movement in its daily life and in extreme cases can result in death. Both parasitic species can move at around 30km per hour and in some cases can sustain this for up to 900km.
The serious threat offered by parasites has a pronounced effect on reindeer movement and herds have developed specific anti-parasite measures. In addition to speed of movement, known to be up to 50km over long distances, they possess special adaptations in the tarsals and metatarsals, which produce a click on movement, and they move in a series of zigzags, often grazing when the opportunity arises. Movement is often into the prevailing wind to dislodge parasites. Such movements, of course, generate greater carbon dioxide exhalation and consequently a greater beacon for parasites. The herd's problem is not actually resolved until they locate cooler, often higher, ground at the end of the summer.
Known predators of reindeer herds, particularly those on the move, are wolves and humans. Wolves are known to isolate juveniles, the old, the sick and single individuals. Individual security lies in the anonymity of the larger herd. Major river crossings are a known hazard to herd movement and many examples exist of considerable loss of life during such a manoeuvre.
Reindeer remains from Pin Hole Cave are itemised and summarised for each stratigraphic level. A total of 1051 bone fragments have been recorded for this study. In addition to those recorded within museum collections, a large number were recovered by the author from the 'store' left by Armstrong within the passage area of the cave. A large proportion of these remains have little or no associated details. Remains that have sufficient dimensional details of their original pre-excavation location are incorporated into a reconstruction of the temporal and spatial distribution within the sediment infill. It has been possible to incorporate 22% of the collection within the main reconstruction and a further 43% are grouped according to stratigraphic level. Under the circumstances, comment is based upon just 24% of the total known bone collection. Antler remains are analysed separately and must therefore be treated with caution.
The large quantity of Reindeer remains occur throughout the cave deposits but appear to concentrate within four groupings as shown below. A total of 2038 antlers were discovered within the cave. The reconstruction incorporates 162 of those finds that have sufficient information.
Area 60-70ft (18-21m) within depths 6-12ft (1.8-3.6m): A total of 118 bones were found within these levels. In addition 108 bones can be located to the passage levels. In effect this means that 38% of the post-cranial bones, less antler remains, occur within the area.
The reconstruction and passage incorporate just under 4% of the remains, which in the circumstances is a very small sample. Within this area, 11 of the antlers are cast and 3 are massacred.
Antler morphology shows that antlers in this concentration include 1 female, 5 juveniles and 13 males.
Area 60-70ft (18-21m) north of the entrance within depths 1-5ft (0.3-1.5m): This area has very few remains and 20 bones are recorded.
The morphology of antlers indicate that these levels contain 10 cast and 4 massacred individual antlers.
Based upon antler morphology, the level has 5 males and 10 juveniles.
Area 32-56ft (10.5-16.8m) within levels 5-6ft (1.5-2m): This area in the front of the cave contained 56 adult bones within the two levels. These appear to have been deposited horizontally and occur at a depth that marks the boundary between Middle and Upper Palaeolithic assemblages. Morphology of antler remain indicate 8 cast antler and 4 from massacred individuals.
Antler morphology indicates 4 males and 2 juveniles.
Area within 33-56ft (9.9-16.8m) north from the cave entrance and within 1-4ft (0.3-1.2m) below datum: This area produced very few bones and 39 were recovered.
The frequency and distribution of Reindeer antler have been described and analysed separately. These remains have also been grouped by age, sex and seasonal casting.
Analysis indicates that Reindeer remains are an important element within the vertebrate population of the cave. Distribution varies and there are at least four localised concentrations (above). The species is also frequent in the cave deposits. Remains are particularly associated with the mound of deposit underneath the roof rupture and associated southern gradient. A second concentration immediately in front to the south of this, occupies part of the passage area. An immediate observation is that the species has obviously been of interest to the Hyaena population, Neanderthal groups and possible European Wolf.
Reindeer population survival
Figure 73: Reindeer (Rangifer tarandus) temporal distribution and survival (source data: Digital archive)
The remains of this species survive in a way that contrasts with virtually all other species known from the deposits (Figure 73). The reconstruction of MNI and the survival of their remains clearly shows that from their first occurrence in the lowest stratigraphic levels there is a steady, sporadic, increase in numbers of individuals throughout the Quaternary.
Reindeer remains are frequent within the deposits and are known from 1051 bone fragments estimated to be derived from 121 individuals. Remains occur in virtually all levels from 1 to 13. Many of the remains are well-preserved (see Figure 74), despite their fragility.
The survival of bones in comparison to the number of skeletal parts in a live population is low, and destruction of their bone remains averages 90%, illustrating massive selection or destruction.
Stratigraphic level 10 has the highest survival of bone fragments (5%) from a live population followed by levels 5-7. Levels 2-3 have just fewer than 3%. Although the variations appear minor, they reflect quite large differences in bone and minimum number of individual's frequency. The reconstruction of distribution also includes an illustration of age classes represented within this group.
The population structure evident from these remains is described in terms of adult and juvenile individuals. This is based upon 20% of the surviving remains. The analysis clearly shows that, despite the small sample number of specimens, 27% are derived from juvenile individuals.
Figure 74: Pin Hole Cave, typical, well-preserved, cast fragment of Reindeer antler from the passage of the cave (Jenkinson 1978b, Image courtesy of Manchester Museum)
Reindeer attrition
The variation of surviving Reindeer body parts is shown in Figure 75, which shows the relatively high survival of head fragments, a low frequency of limbs and very few axial parts. Head parts are grouped as a total but many of these are antlers or antler fragments, some of which are cut, polished or engraved. The high number of antlers includes many that are naturally cast with very few massacred examples.
Figure 75: Reindeer (Rangifer tarandus) body part selection (source data: Digital archive)
The high frequency of occurrence also suggest that calculations of a minimum number of individuals, which are largely based upon the smaller numbers of post-cranial bones, may under-represent the actual numbers that may have been present within a live population from which such a selection was made. The combination of these observations suggest a clear indication of human agency, which contrasts with that observed for carnivores. Limb parts are relatively frequent among the remains but there are very few axial parts.
The evidence available from body parts demonstrates the selective use or collection of heads, mainly cast antlers and some limb bones, both of which are transported into the cave. This suggests that human groups were collecting antler as a raw material in conjunction with limbs as a food resource.
Figure 76: Reindeer (Rangifer tarandus), attrition, bone damage (source data: Digital archive)
The degree of damage to this species is graphically shown in Figure 76. A very high frequency of the surviving bone is fractured but is also associated with a reasonable number of complete bones.
Gnawed bone occurs in stratigraphic levels 11 and 12, where Hyaena residence is known. A relatively high and abnormal quantity of gnawed pieces occur within the upper levels (1 to 3) which, given the relative absence of Hyaena in these levels, were probably the result of Wolf activity. The overall temporal trend of bone damage is remarkably similar to that recorded for Bison. In both instances, the coincidence of clustering, particularly of fractured and complete undamaged bone within levels with archaeological remains suggest that human agency is a principal cause. Gnawing of bone is recorded within stratigraphic levels 11 and 12, but its occurrence is interrupted suggesting that carnivore, specifically Hyaena, populations may have been periodically absent or may pre-date human use. In a similar fashion, gnawing within the upper levels is interrupted and its occurrence appears to decline within each identifiable cluster where human agency is known. In this instance, the degree of overlap in the occurrence of gnawing with other forms of bone damage reflects the lack of fine definition within the sequence. The apparent coeval evidence for humans and European Wolf is intriguing, particularly as much of the gnawing on some bones is probably the result of Wolf activity.
Temporal and spatial distribution of Reindeer antler
Antlers and antler fragments are plentiful within the cave and 2038 specimens are incorporated into the Digital archive. Sufficient dimensional details exists to incorporate 162 within the main reconstruction and to group a further 38 into a passage context. In effect, 17% of antlers can be sourced to the deposits.
Concentrations of antler within the illustrated distribution are difficult to perceive, particularly in the context of such a small sample. There is some evidence of a rear concentration with the area 51-73ft (15.5-22.2m) north of the entrance and within depth levels 8-12. This concentration appears to slope to the south. A second cluster is evident with the area 12-60ft (3.6-18.2m) north of the entrance and within depth levels 104ft (31.6m). This concentration seems to have a horizontal distribution.
Antlers are grouped within the analysis to describe both sex and casting state. There are low numbers of 'massacred' antler and their distribution appears to concentrate in the rear of the cave. A very large proportion of antlers are cast and from juveniles. This is of particular interest in that it indicates a presence in summer and, in all probability, late summer and autumn. In terms of human groups, this antler must have been collected locally and stored in the cave as a resource. There are multiple examples of both finished items and cut and chopped antler sections known from the cave, particularly in the Later Palaeolithic. A small number of pieces are gnawed by carnivores in a fashion that is distinctive to European Wolf. This is likely to be coeval with the year of casting when the antler has a remnant of fleshy content.
Reindeer incised and cut-marked bone
Figure 77: Drawing of Reindeer (Rangifer tarandus) antler fragment showing surface cut-marks and 'snapped' distal end (Drawing by Andy Palmer)
The upper stratigraphic levels, which are associated with Later Upper Palaeolithic artefact assemblages, also contain numerous cut or incised fragments of Reindeer (Rangifer tarandus) antler. The example shown in Figure 77 is of a juvenile female cast antler and has a sharp oblique fracture at its lateral end. The surface has three distinct 'chop' marks, all on the same side. The lateral fracture is possibly due to attempts to snap the antler and certainly there is no evidence of carnivore surface damage. It is interesting to note that the distance between chop marks is of the same size order as on pieces of decorated antler known from the cave and may therefore be good evidence for the preparation of antler for engraving.
6.6.8 Giant Deer (Megaloceras giganteus)
This species is the largest of the deer genera known from Quaternary deposits and is thought to have an ethology similar to modern Elk (Alces alces). Although remains are not common, the species is known from the Middle Quaternary onwards. Modern Elk populations show a preference for open woodland with plentiful water resources. Body length for this species is usually between 2.5 to 3m, shoulder height between 1.3 to 1.9m and with a body weight of approximately 825kg (Walker et al. 1975). Individuals are solitary or travel in small groups and can move at a speed of 25 to 35km per hour. They are browsers and have a wide dietary preference, which includes trees, shrubs and water plants. Identification of Pin Hole bone remains and particularly distinguishing them from Red Deer (Cervus elaphus) is extremely difficult for all skeletal parts except antlers. Within this study, analysis of the remains show that they share many common morphological traits, suggesting they all belong to one species. Bearing in mind the size, particularly of dentitions, the material has been referred to the species Megaloceros.
Later studies of the material by Currant and Jacobi (2001) have referred the remains to the species Red Deer (Cervus elaphus). Modern study, using DNA bone collagen fingerprinting, has been unable to distinguish between the two species. It is clear that the palaeontological remains belong to one large Cervid animal but it remains uncertain which species they are derived from. Giant Deer remains are itemised in the digital archive and also summarised for each stratigraphic level.
Giant Deer Population survival
Figure 78: Giant deer (Megaloceros giganteus) temporal distribution and survival (source data: Digital archive)
The remains of Giant Deer are known from 19 bone fragments estimated to be derived from 9 individuals (Figure 78). The species are represented by a very low frequency of bone fragments, often the remains of single individuals within separate levels. Temporal distribution is primarily concentrated within stratigraphic levels 6 to 12 with occasional occurrence in levels 4 and 2. Bone survival of the live population is poor and clearly indicates selection of particular skeletal parts or their destruction by carnivores. Correlation with archaeological data show that the main occurrence is within levels containing evidence of the Middle Palaeolithic and Hyaena populations.
Giant Deer attrition
Figure 79: Giant Deer (Megaloceros giganteus), body part selection (source data: Digital archive)
Despite the low number of individuals, the dominance of head parts associated with limb fragments (Figure 79) and an absence of axial parts suggest predation, where individual or inter-species competition at kill sites results in the removal of accessible parts consumed or cached elsewhere. In such circumstances, it is likely that axial parts were never introduced into the cave. The identity of the predator is not clear and the overall low number of individuals suggests that this is an occasional food resource used sporadically or opportunistically.
Giant Deer attrition, bone damage
Figure 80: Giant Deer (Megaloceros giganteus), attrition, bone damage (source data: Digital archive)
Many of the species' bones are fractured and these are associated with bone that is both fractured and gnawed (Figure 80). The mandible fragment in Figure 81 shows typical Hyaena fracturing, with lateral bite marks and surface gnawing.
Figure 81: Fractured and gnawed mandible of Giant Deer (Jenkinson 1984; Image reproduced by courtesy of Manchester Museum)
The most likely cause of the species' introduction into the lower levels is that of Hyaena predation activity where body parts have been cached or introduced by breeding groups. The presence of gnawed parts suggests that juvenile Hyaena were present. Stratigraphically, the remains are in levels that contain abundant evidence of Middle Palaeolithic use of the cave but the style of body part selection, the type of damage and the lack of any evidence of fracturing or cutting by human groups supports the view that Hyaena were the main agency of accumulation.
6.7 Hares and Ground Squirrels (Lagomorpha)
6.7.1 Ground Squirrel (Spermophilus major)
Modern Ground Squirrels are known from 14 species and are present throughout America and Asia. The species are adapted for ground living and prefer burrows, hollow logs and rocky areas. Their diet includes seeds, nuts, plant stems, mice, insects, birds and eggs. Food is temporarily stored in cheek teeth and permanently underground. Populations are known to prefer arid and grassy prairies and rocky areas (Walker et al. 1975).
Three specimens of this genera have been identified to Spermophilus. The taxonomy of 14 modern Spermophilus species is confused, but this study has followed the comments of Mayhew (1955).
A left mandible has been identified to the species. The mandible is labelled by Armstrong as derived from depth level 9-10ft (2.7-3m) within the 'Inner Chamber' and associated with the 'M2' middle Palaeolithic. This is identified to Spermophilus major following Mayhew (1955). A second specimen, a matching mandible from the same individual, which has no dentition, is recorded from 'D1' 1ft (0.3m depth) in the rear of the cave. A third specimen, a tibia shaft, is recorded from 66-70ft (18-21m) north from the entrance at a depth of 6ft (1.8m). This specimen is within the same size category as the previous two specimens but it is not possible to affirm that it is from the same individual. The distribution of the three specimens is illustrated in the digital archive.
Despite there being only three specimens, they are discussed in detail as they are rare in the UK record and attention has been drawn to the importance of these specimens. Later studies by Currant and Jacobi (2001) report confirmation of the species' identification and include them as a component of the MAZ 'Mammoth Steppe' fauna.
There is a difficulty concerning the location of these specimens. The two mandibles are without doubt from the same individual. One is from the 9-10ft (2.7-3m) level in the rear of the cave while the matching mandible half was discovered within the same general area at a depth of 1ft (0.3m). The third specimen is from the same area but from 6ft (1.8m) depth. Although the last specimen cannot be directly related to the first two, it is clear that they are derived from the top of the sedimentary section. The circumstances are complex as the left mandible, which is the one identified to species level, is from the 'inner chamber' at 10ft (3m). This is exactly the location where the later glacial sediment and flowstone has formed down the eastern cave wall from the top of the section and is known from 1986 excavations to have filled parts of the 'inner chamber'. Under the circumstances, the specimens are of uncertain age but cannot be considered to belong to the lower sediments that are suggested to represent MAZ (Mammal Assemblage Zone).
6.7.2 European Hare (Lepus timidus)
Hares are primarily woodland species that reside in burrows. They display a modern-day distribution that is very diverse and incorporates a very wide range of habitats (van Den Brink 1977). A total of 62 bones were identified to the species and these are illustrated within the stratigraphic distribution shown in Figure 82.
Figure 82: European Hare (Lepus timidus) temporal distribution and survival (source data: Digital archive)
The distribution of surviving remains of this species show that only a small quantity of the original live population skeletal remains have survived (Figure 82). There are few remains of the species in the lower deposits, most of which occur from the surface to stratigraphic level 6. Comparison of surviving bones from a live population indicated that over 80% are destroyed or missing. The distribution of remains correlates closely with that for both Red Fox and European Wolf (see Figure 45, Figure 32), which suggests predation by these carnivores.
European Hare attrition, body parts
Figure 83: European Hare (Lepus timidus), body part selection (source data: Digital archive)
Hare body part selection is dominated by cranial and limb bones, which represent over 70% of the surviving remains (Figure 83). Axial body parts are present in low numbers. This suggests that the whole carcass was originally introduced into the cave and where overall destruction or consumption of the remains has taken place within the cave interior. The low incidence of axial parts, particularly vertebrae, may be an artefact of the analysis in that these may be present within collections of microvertebrate bones that have not been identified to this species.
Bone damage and attrition is illustrated in Figure 84. The figure clearly shows that Lepus bone survives in a complete or fractured state with little evidence of gnawing or bite marks. No evidence was found of artefact cut-marks.
Figure 84: European Hare (Lepus timidus), attrition, bone damage (source data: Digital archive)
The stratigraphic correlation between this species and the Canids (Red Fox and Wolf), the pronounced selection of axial body parts, the survival of limbs and crania and the absence of cut-marks clearly suggest that the species has been subject to Canid predation or the remains were the result of natural casualties.
6.8 Rodent genera and species
A wide range of Rodent species are known from the cave (Jackson 1934a, 1934b), and these have been identified mainly upon the morphology of crania, mandibles and isolated dentitions.
Of the total Rodent bones from Armstrong's excavation, some 165 have been identified to species whilst 3944 are identified to genera only. This clearly presents some difficulty, in that the analysis of species alone would represent a small quantity of the total known remains. In each of the figures shown below, both species and genera are used and the two species of Lemming are considered individually. Before considering the analysis, it is worth noting the fact that Leslie Armstrong managed to recover and record such micro remains in quantity from such a difficult excavation environment.
The analysis illustrated below is also based upon morphological variation in bone parts to aid species identification. Modern studies, utilising the DNA collagen finger printing are currently being undertaken (Buckley et al. 2017) and it is highly likely that the results of such analysis will alter the details of the known species and their distribution used below. It has also been suggested (Jacobi et al. 1998) that remains of these species, owing to their size and burrowing behaviour, may well have become mixed or incorporated within earlier stratigraphic levels. There is no direct evidence from Armstrong's excavation that suggest movement on a significant scale. Later excavation by Jenkinson in the rear of the cave coupled with detailed statistical studies of the fauna (Ringrose 1990) suggests that little movement of remains has taken place. Such movement is also seriously inhibited by the large amount of limestone clast within the sediment, making burrowing difficult. Additionally there are very few examples of modern populations resident in the dark rear of caves. It is simply not appropriate to use the behavioural patterns of modern rodent populations as a means of explaining away confusion within the stratigraphic interpretation of other types of data.
Figure 85: Rodent species temporal distribution (excluding Lemmings) (source data: Digital archive)
The nature of the data restricts realistic comment. The analysis show in Figure 85 is based upon less than 5% of the fossil record that has been identified to species. The analysis simply shows that these species seem to have had a consistent presence within the cave throughout most of its history. The occurrence of Hare in Levels 1-5 replicates that seen for many other species, particularly the megafauna. Stratigraphic levels 11-12 have an isolated cluster that includes four species of Vole associated with Lemmings. The level above contains evidence of the only Ground Squirrel known from the Creswell sites and shows an increase in numbers and a diversification of species seen for many insectivores, birds, amphibians and fish known from the cave.
Figure 86: Rodent species and genera, bone distribution, body parts and survival (source data: Digital archive)
The data illustrated in Figure 86 includes all of the genera Rodentia recovered from Armstrong's excavations that have sufficient stratigraphic information to reconstruct their original stratigraphic location. Rodentia are represented by relatively large numbers of bones (and MNI) but do not survive well. Armstrong's excavation methods recovered large numbers of bones of diverse skeletal type so it would seem that the extant collection is a real reflection of the original circumstance. Rodent is present within most stratigraphic levels. The figure represents survival and MNI based upon cranial remains and excludes a large amount of post-cranial material; nevertheless, the survival rate for species appears higher than usual for the vertebrate remains within the cave.
Figure 87: Rodent species and genera, body part distribution (source data: Digital archive)
Figure 87 attempts to show a more realistic distribution of Rodent remains by including the large quantity of bones that have been identified to generic level within their stratigraphic context. The analysis shows that a large number of limb bones are present with very few associated axial bones; these are often scapula and pelvic fragments. Vertebrae remains are rare. This circumstance could be the result of differential recovery during excavation, which despite excellent recovery of micro remains may not have recovered very small vertebral fragments. In general terms, this type of preservation is well known from numerous studies of the regurgitated pellets of raptors and small carnivores, where prey is often swallowed and digested whole, without chewing. In many predatory species, the larger bones and fur are regurgitated, often in specific well-used locations that over time form accumulations.
Figure 88: Rodent genera, attrition damage (source data: Digital archive)
Analysis of damage to Rodent bone clearly shows that skeletal remains are either complete or damaged by simple fractures (Figure 88). There is no evidence of gnawing. Analysis also suggests that many fractures may be post-mortem, particularly as the respective items have often been found closely associated within the sediment. Despite detailed searching, analysis of the remains did not reveal any sign of bite or killing marks (typically inflicted by raptors, especially Owls). This also suggests that many species either died in situ as residents or were prey that was swallowed and regurgitated whole. There is no specific recording from Armstrong concerning accumulations within the cave, with the single exception of his so called 'rodent corner'. It was never clearly stated where this was located but it seems likely that it was at the junction of the passage and the 'inner chamber' along the eastern wall.
6.8.1 Lemmings (Microtidae)
Arctic Lemming (Dicrostonyx torquatatus) and Norway Lemming (Lemmus lemmus) are known from the cave by the presence of 1233 identified parts, mainly cranial or isolated dentition. Both species are nocturnal and occupy open grassland habitats. Modern populations migrate into both birch and conifer woodlands. Breeding takes place in spring, when nests of moss and grass are constructed on the ground surface. Modern populations are preyed upon by a wide range of species, which include Wolf, Fox and a range of raptor birds (van Den Brink 1977).
Arctic Lemming and Norway Lemming have been identified using dental fragments, which are among a very large collection of post-cranial Rodent skeletal parts known from the excavation. Sufficient dimensional detail is present to allow incorporation of 332 dental parts within the reconstruction and a further 43 can be located to the passage levels. The reconstructed remains are 78% of those identified but are less than 1% of the known Rodent bones. Despite being a prolific body of evidence from the cave, the lack of dimensional information for many specimens very seriously inhibits any interpretation of their relevance. Comment therefore cautiously considers location to be particularly within the lower deposits. The status of Rodent remains and Lemming species in particular is one of the very difficult aspects of evidence from Pin Hole. Their frequency, distribution and lack of reliable identification combine to render the data almost useless at present.
Figure 89: Arctic Lemming (Dicrostonyx torquatus) and Norway Lemming (Lemmus lemmus) temporal distribution and survival (source data: Digital archive)
The stratigraphic distribution for both species (Figure 89) is principally within the lower sequence (levels 10 to 13), with more minor peaks of occurrence with levels 5 to 7 and 1 to 3. The fluctuating occurrence of both species does not seem be directly related to fluctuations in carnivore species, which suggests the availability of preferred habitat conditions. Survival of skeletal remains is unusually good. The varying density may reflect temporal change in access to sub-arctic moorland or open ground. Surprisingly, the distribution correlates well with that of both human use and raptor presence within the cave.
6.9 Insectivores and Bats (Insectivora and Chiroptera)
Chiroptera or Bat remains have been extensively studied by Dr Robert Stebbings for all of the Creswell localities and are partially reported in Jenkinson (1984; Jenkinson and Gilbertson 1984). The remains are described here for the first time.
A total of 10 bat species have been identified and a further 264 specimens have not been subject to detailed study (Figure 90). All of the species are insectivores but have a fairly wide range of habitats that include woodland with open spaces and watercourses. The presence of so many species within such a small site appears to show a high degree of naturalism between species with little indication of intra-species competition.
Largely complete, undamaged bones represent individual species from all body regions. The cave environment offers a wide range of rock crevices, ledges and holes and it is probable that these remains represent fatalities among winter-hibernating or spring-breeding colonies. Their sudden appearance in quantity within stratigraphic levels 0 to 4 indicates the presence of woodland and watercourses with large insect populations, especially locally, within the limestone gorge itself. All of the species are absent from the lower sequence between stratigraphic levels 7-20 (where the species that are found - ground-living mammals, birds and amphibians - preferred open ground with scrub but lacking woodland).
Figure 90: Bat species (Chiroptera) temporal distribution (source data: Digital archive)
More bats are known from Pin Hole than the total from all of the other caves. This circumstance may be due to its open aspect and position relative to adjacent watercourses and open areas.
6.10 Birds
6.10.1 Ducks and Geese (Anatidae and Anseriformes)
The extensive collection of avian remains has only been identified in recent years and is known almost exclusively from the research of Dr Don Bramwell (Jenkinson and Bramwell 1984). The results are published in part in Jenkinson and Gilbertson (1984) and a more comprehensive list was published in Jenkinson (1984). Further studies have reported duck, geese and egg fragments (Jenkinson and Sutherland 1984) and their importance as indicators in change of migratory behaviour. Other studies have re-examined identifications for some avian genera (Stewart and Jacobi 2015). The known collection has been inventoried and described in the digital archive).
Temporal and spatial distribution of Ducks from Pin Hole Cave
Eight species of Duck have been identified from a total collection of 156 bones and 58 of these have sufficient dimensional information to be incorporated into the reconstruction (Digital archive). This represents approximately one quarter of the known specimens. The remainder are not yet identified. Identified species include Common Scoter (Melanitta nigra), Mallard (Anas platyrhynchos), Ruddy Shelduck (Tadorna ferruginea), Tufted Duck (Aythya fuligula), Widgeon (Anas penelope), and marine-coastal species Red-Breasted Merganser (Mergus serrator), Goosander (Mergus merganser) and Garganey (Anas querquedula). The remains also include egg-shell fragments and at least one group of complete eggs (Figure 95).
Most of the remains are within depth levels 1-5ft (0.3-1.5m) in an area within 30-60ft (9-18m) north from the entrance. The distribution is random within this area. A second cluster occurs between 50-71ft (15-21m) north of the entrance and within levels 9-12ft (2.7-3.6m) below the datum. This group includes Pink Footed Goose.
Temporal and spatial distribution of Geese from Pin Hole Cave
Six species of geese have been identified from of 330 bones. A total of 90 have sufficient dimensional information to be incorporated within the reconstruction. The species include Greylag Goose (Anser anser), Barnacle Goose (Branta leucopsis), Brent Goose (Branta bernicla), Pink-Footed Goose (Anser brachyrhynchus), White-Fronted Goose (Anser albifrons) and Bewick's Swan (Cygnus columbianus (Digital archive). There are a number of egg fragments associated with the remains.
The temporal and spatial distribution is very similar to that recorded for ducks in that most of the remains are within the upper 5 levels. There is a second group of remains within depth levels 9-12ft (2.7-3.6m) below datum within the area 50-71ft (15-21m) north of the entrance. The distribution of Geese is shown in Figure 91. There is also a large sample of avian bone from the excavation that has not been identified and which may contain additional evidence of other species.
Figure 91: Temporal Distribution of Geese Species (Source data: Digital archive)
Both Duck and Goose genera have similar stratigraphic distributions. Geese species (Figure 91) have a clumped distribution and are concentrated within levels 8 to 11 and 2 to 5. A rather obvious point is that this coincides with the distribution of egg fragments and which indicate the various species' presence during the early part of the breeding season, within late winter or early spring. All of the species are relatively large, form large flocks and are known to migrate into breeding areas (see Jenkinson and Sutherland 1984). The lack of juvenile individuals preserved within the cave seems to indicate an absence of the genera in late summer and winter. Their occurrence also indicates the nearby presence of extensive areas of water and open shallow aquatic habitats which provide appropriate grazing.
Very low numbers of Goose species are recorded from stratigraphic levels 8 to 12, levels 5 and 2 to 3. The remains of one or two individuals, or four individuals in the case of Pink Footed Goose, Bearing in mind the numbers that were probably present within summering flocks, the remains seem to form a very rare component of the vertebrate population. Analysis of bone body parts present and the attrition damage shows that the few individuals are represented by limb parts, particularly coracoid or wing pieces with no axial parts (Figure 92).
Figure 92: Goose humerus and coracoid from Pin Hole Cave (Reproduced courtesy Manchester Museum)
All six species prefer open areas, particularly with shallow freshwater. Their occurrence is sporadic. They are often represented by single skeletal parts and less than 1% of the original bird skeleton appears to have survived. They occur, often as single bones, in all stratigraphic levels with the exception of level 7.
Duck and Geese species attrition
Figure 93: Duck and Geese, attrition, bone damage (source data: Digital archive)
Bone attrition for duck and goose species are grouped by genera and the distribution is shown in Figure 93. Virtually all of the duck and geese skeletal parts are complete (44 complete, 3 fractured). Examination of the few body parts shows that virtually all are limb bones. Recorded attrition patterns are of a distinct type and suggest that whole or near whole carcasses have been transported into the cave where both cranial and axial sections of the body have been consumed and where the heavily feather-covered wing parts, of little food value, are waste.
This pattern of survival suggests that they represent a form of predation where scavenging or kill may have taken place outside of the cave environment and wing parts only are transported into the cave interior. It is probable that this represents predation by small carnivores, particularly Red Fox or European Wolf, both of which are known within the levels where this evidence occurs.
Figure 94: Goose, coracoid and humerus shaft showing two bite marks near the distal epiphysis and a bite mark that has fractured the proximal epiphysis (Image courtesy of Manchester Museum)
Direct evidence of the taphonomic circumstance of accumulation is available from at least one set of Goose remains found within stratigraphic level 2 (Figure 94). The lower of the two illustrated bones shows a damaged Anseriform humerus where two very clear canine impressions are present on the distal shaft. The proximal shaft on the same piece bears a very distinctive fracture resulting from carnivore puncture of the bone shaft within this area. The size of the canine marks on the bone surface suggest that they are of a species of cat (Felidae sp), Red Fox (Vulpes vulpes), or small Mustelid, all of which are known within these levels.
Anseriform egg fragments
Figure 95: An example of a complete egg (one of several) found within the cave during Leslie Armstrong's excavation (Jenkinson 1978b; Image courtesy of Manchester Museum)
Several substantial parts of Anseriform eggs were unearthed during excavation within the passage of the cave, and at least one group of 7 eggs contained complete examples (Figure 95). It is not known how many were complete on discovery but Armstrong reports one group of 6. An additional report in 1927 (Garrod 1927) refers to the discovery of four large eggs. The egg shown in Figure 95 has been subsequently filled with fine-grain sediment and the surface infill indicates the original state of the eggshell. This shows a contemporary fracture in the egg surface where contemporary sediment protrudes (top right of photograph). This may show an original puncture within the egg's outer case, which could be due to a carnivore puncture. Whatever the circumstances of collection and transport into the cave, it is a remarkable piece of evidence, which offers comment not only on the circumstance and care exercised by a carnivore or possibly a human group, but also on the nature of the dynamics of sedimentation and the low-energy nature of movement within the deposits.
There is a general distribution of shell fragments in many stratigraphic levels (Figure 96). These record the distribution of 153 shell fragments from at least 7 stratigraphic levels of the cave.
Figure 96: Stratigraphic distribution of Anseriform egg fragments (source data: Digital archive)
The distribution of eggs is distinct, with a concentration in level 10 and within level 12; the location of the undisturbed group and a more numerous distribution in the upper sequence represented by levels 1 to 6 (0.3-1.8m). There are several instances when eggs were discovered complete and unbroken. In one instance, Armstrong notes that a flint blade was found next to egg fragments.
The upper levels have large numbers of both Wolf and other smaller carnivores such as Red Fox, Marten and Weasels, all of which may have been responsible for egg accumulation and fragmentation. The occurrence of a group of eggs within level 12 is more difficult to explain and it is possible that the species responsible was nesting in the then open and near-empty cave passage. It is also possible that Hyaena or European Brown Bears may be responsible; both are known from modern studies to collect and cache such resources.
Raptors are a rare discovery within this locality and a total of 78 bones are known, which have been identified to five Falcons and five Owl species. Twenty-seven of the bones have sufficient dimensional details to be incorporated within the reconstruction. Virtually all of the specimens were found within depth levels 1-4ft (0.3-1.2m). This applies the whole length of the cave. Osprey and Barn Owl are known from single specimens at 60ft (18.2m) north of the entrance and within the 8-9ft (2.4-2.7m) levels.
Falcon and Eagle remains are illustrated in Figure 97, which shows the six species have been identified primarily from stratigraphic level 0-3. Lower levels have evidence of Golden Eagle (Aquila chrysaetos), Rough Legged Buzzard (Buteo lagopus), Merlin (Falco columbarius) (level 6) and Osprey (Pandion haliaetus) (level 11). Kestrel (Falco tinnunclus), which is a resident of the site today, is found throughout the whole sequence. With the exception of Kestrel all other species are represented by the remains of one individual. The survival of bone from a live population of these species is exceptionally low. For most species less than 1% survives. Stratigraphic levels 1-3 exhibit slightly better survival but even within these levels only 3% survive.
Modern populations of these species are resident within a wide range of habitats, which include open ground and woodland or a combination of both. Osprey is a migratory species and often associated with the presence of aquatic environments and open ground.
Figure 97: Eagle and Falcon species (Falconiformes) distribution (source data: Digital archive)
As a result of the small size of the overall sample, a detailed analysis of their skeletal part variation and attrition has not been undertaken. It is, however, clear those fragments of limb bones represent most species and that most of these remains are complete or have simple fractures.
For many of the larger species, access and possible residence within the narrow cave is very unlikely, due not only to the restrictions within the cave but also the lack of potential perching areas, which are only present within the cave entrance and adjacent rock faces. Despite the careful nature of Armstrong's excavation and his recovery of prolific micro-faunal remains, there is no extant evidence of pellet remains, which might have suggested residence. Under the circumstances it seems very likely that small predators who have selected or destroyed the central axial and more edible body parts have occasionally scavenged the species.
The stratigraphic distribution of species correlates closely with that of Rodent species, Grouse (from Buzzard diet) and fish (from Osprey diet). Their presence and gradual increase in frequency within the upper section of the cave seem to indicate nearby open ground with tree cover and water sources.
6.10.3 Owls (Strigiforms)
Five Owl species are known from stratigraphic levels 0-2. They are all small species and include Tengmalm's Owl (Aegolius funereus), Short Eared Owl (Asio flammeus), Tawny Owl (Strix aluco), Barn Owl (Tyto alba) and Hawk (Surnia ulula). Most of these species are first records for a UK cave site and these are shown in Figure 98.
In common with the remains of raptors, it is unlikely that they were resident due to the caves restricted nature and the lack of perching areas within the main cave. They may have utilised the extensive rock ledges and solution holes immediately above the cave entrance and adjoining rock faces. The few remains discovered are primarily limb bones and tarsals, most of which are complete.
Figure 98: Owl species (Strigiformes) distribution (source data: Digital archive)
There is no tangible evidence of a taphonomic process explaining their presence within the site but their rarity and lack of cranial and axial parts suggest that small carnivores might have scavenged them. The lack of evidence for pellet remains, which would indicate residence, seems to support such a view. Pellet remains may exist adjacent to the prominent rock faces adjoining the cave entrance. There has been no systematic excavation of this area. Records of such activity have been recorded within modern populations.
The occurrence of all four species, which prefer open woodland habitats and have a varied diet of rodents, small bird species, amphibians and insects, also suggest that such a habitat existed within the immediate area of the Creswell gorge and adjacent valleys.
6.10.4 Grouse (Galliforms)
Grouse species present within the area today are confined to Partridge; all other species are locally extinct. Ptarmigan (Lagopus muta) usually occur above the tree line at 600m or more. Red Grouse (Lagopus lagopus) and Black Grouse (Tetrao tetrix) normally prefer open landscape with restricted tree cover, where their main diet is heather.
Galliform genera are known from 505 bone fragments, which have been identified to include five species and Grouse genera. Sufficient dimensional details are present on 130 specimens and these are included in the reconstruction. This represents 15% of the known specimens. There are two main concentrations of bone. The first occurs within the area 47-66ft (14-19.8m) from the entrance within depth 3-7ft (0.9-2.1m). This concentration is within the mound of sediment at the cave rear and the distribution appears to show evidence of the incline toward the south and entrance. The second concentration is more general and extends laterally and horizontally within 33-75ft (10-22.8m) from the entrance and within the upper levels.
Figure 99: Grouse species and genera (Galliformes) temporal distribution (source data: Digital archive)
A total of 73 bones have been identified to four species and 19 bones to the genera (Figure 99). Stratigraphic levels 7-12 contain Ptarmigan and Red Grouse. The remains of the estimated seven individuals survive poorly, between 1-2% of the original skeleton. The upper sequences (stratigraphic levels 0-6) contain all 4 species and genera. The survival of each species' remains, while very low percentages of the original live skeleton, averages 3-7%. This is a clear indication of a considerable change in the taphonomic circumstances of their accumulation.
Figure 100: Grouse genera and species, body part selection (source data: Digital archive)
Body part survival for all species and genera of Grouse are shown in Figure 100. Skeletal remains are dominated by limb bones The results replicate the pattern of survival already discussed. The lower sequence 7-12 is dominated by limb parts with an unusual survival of axial parts in Level 7.The upper sequence 0-6 is also dominated by limb parts but has both cranial and axial parts.
Damage and attrition to Grouse bone is shown in Figure 101. The chart clearly shows that practically all of the bone is either complete or has suffered fracture damage and that this is associated with individual specimens which have either puncture or gnaw marks (see Digital archive).
Figure 101: Grouse genera and species, attrition, bone damage (source data: Digital archive)
The stratigraphic distribution of Grouse correlated very closely with that of European Wolf and, to a lesser extent, with Red Fox. The nature of body part selection and individual bone damage is typical of the predation behaviours of both species in that the smaller and axial parts of prey are consumed and the less meaty limb or wing parts are often damaged but not swallowed. It has been suggested (Campbell 1977) that these genera may have been a food resource for human groups. This may be a possibility as this analysis shows that the stratigraphic and temporal distribution closely follows that of human use of the cave. Unfortunately, it is also mirrored by the distribution and increase in Red Fox, European Wolf and Raptors.
Deposition within the cave sediments may be due to several agencies. The only direct evidence available are the small carnivore bite marks on some bones (Figure 94). Many of the specimens were examined for signs of human fracturing and cutting marks, but none was found. There is insufficient evidence but at least two of the concentrations appear to have distributions associated with the later Middle Palaeolithic and Upper Palaeolithic.
6.10.5 Waders (Ciconiiformes and Charadriiformes)
Species from both Cranes (Ciconiiformes) and Waders (Charadriiformes) families are included within this group as all of the species obtain food by wading in various depths of water. The groups have a modern Holarctic range and breed during the summer in low tundra environments with adequate supplies of moss and aquatic plants and animals (Soothill and Soothill 1982). Two species, Lapwing (Vanellus vanellus) and Plover (Pluviallis squatarola), are seasonally present on the site today. The remains from Armstrong's excavations are described in the digital archive.
The fifteen species of Waders known from this excavation are shown in Figure 102. Wading birds have been recovered from various levels within the cave. A total of 45 bones are known, 39 of which have sufficient stratigraphic details to be included in the reconstruction. The range of species is of some interest. Demoiselle Crane (Anthropoides virgo) is a unique British occurrence and is known from a depth of 3-4ft in the 'inner chamber' (0.9-1.2m). Other very unusual occurrences include Whimbrel (Numenius phaeopus), Redshank (Tringa totanus), Greenshank (Tringa nebularia), Snipe (Gallinago gallinago), Turnstone (Arenaria interpres), Curlew (Numenius arquata) White Stork (Ciconia ciconia) and Plovers (Pluvialis).
Figure 102: Wader species (Ciconiiformes and Charadriiformes) temporal distribution (source data: Digital archive)
The species are represented by bones in a complete state. The specimens are of adult birds and there is no evidence of juvenile material or egg fragments. Despite the extremely small sample it is possible to infer that they represent an occasional occurrence of a summer population that has access to non-frozen water supplies and associated resources
The species are migratory over large distances. Crane and Stork species currently winter in regions to the south-east, particularly within the Black Sea area. The occurrence of Demoiselle Crane is very unusual. The species currently spend summers in eastern steppes and North India. Many of the Charadriiformes currently migrate over long distances to mid- and West African regions. The occurrence of these groups therefore offers direct evidence of a recent northerly shift in home range (see Jenkinson and Sutherland 1984). It is difficult in such a small sample to infer taphonomic factors that may have affected their accumulation within the cave. However their presence does indicate that aquatic food resources, mosses and low-growing herbs within a river and lacustrine, low tundra and open grassland, environment are locally or regionally present. Several of the larger species hunt and consume rodents, reptiles and frogs. Several of the species were resident in the waterlogged areas that existed to the south of the cave until quite recently. The presence of many of the species clearly indicates that on several occasions the local region contained a diverse population of aquatic and terrestrial invertebrates for which there is currently no other evidence.
6.10.6 Maritime species - Auks (Alcidae)
Nine bones have been identified to four species of Auk, which are unusual, and of considerable interest (see: Digital archive), Gooseander (Mergus merganser), Gull (Larus canus), Black Guillemot (Cepphus grylle) and Puffin (Fratercula arctica). With the exception of Gull, none of the species is regionally present today. Merganser, a diving duck, is included within this group due to its current coastal distribution. Modern populations prefer coastal areas, particularly those with rocky ledges and cliffs and all species spend the winter season at sea and use a land location for breeding.
The sample is too small to realistically consider the taphonomic circumstance surrounding their incorporation within the cave deposits but their presence is likely to be the result of regionally scavenged remains. It seems likely that they indicate summer presence in regional coastal areas. The intermittent occurrence of the species within stratigraphic levels 0-6 does at least indicates the local existence of a maritime and coastal habitat.
6.10.7 Woodland bird species distribution
Woodland species included within the study include 48 species represented by 1150 avian bones, many of which were identified by Dr Don Bramwell. A further 648 avian bones have not yet been identified and are not included within Figure 103. The remains are described in the digital archive.
Figure 103: Woodland bird species (Passeriformes), temporal distribution (source data: Digital archive)
The total population shown within Figure 103 has been replotted according to the reconstructed stratigraphic levels. The avian record is perhaps the richest from a Quaternary site that is available within the UK. The figure shows that very few remains are found below stratigraphic level 8. Above this layer there is a huge increase in quantity and diversity of Passerine, woodland species.
The dramatic rise coincides with the occurrence and increase illustrated for Owls, Falcons, Wolf and a range of small carnivores and the noted decline in frequency of large carnivores. Their distribution also indicates a presence of woodland locally, which in the uppermost sequence becomes the principal habitat. Species also indicate that other habitats were nearby that included water resources (e.g. Kingfisher), open ground (Buntings), pine woodland (Grosbeaks), and developed ground cover (Finches and Tits). Some of the areas may have been mature woodland (Rooks, Crows and Ravens).
Many of the faunal remains are complete and show little sign of damage by carnivores. There are no reported accumulations or pellets from within the cave, which seems to indicate that predation by medium and small carnivores is the most likely agency of accumulation.
6.11 Amphibians (Frogs and Toads)
Both Common Frog and Common Toad prefer dry to moist areas and are mainly nocturnal. Common Frog is known from high altitudes but not within permanently frozen areas. Both hibernate during winter and congregate in large numbers in rivers and streams for breeding during the summer. Predation of both species is extensive, particularly by birds and small carnivores. Individuals are often swallowed whole. Extensive studies have shown that their bones can be preserved within raptor and carnivore pellets and scats.
The Armstrong excavated collection contains 3192 Common Frog bones, identified by the author in 1977. Of these 1924 have sufficient dimensional details to be incorporated within the reconstruction. In addition, 40 bones can be sourced to the depth levels in the passage. In effect this means that 61% of the excavated collection can be relocated to the cave deposits.
Figure 104: Amphibian survival and body part (source data: Digital archive)
The evidence from the cave is distinct but of low frequency and contrasts markedly with localities where small carnivore predation is present, such as Steetley Cave (Jenkinson and Gilbertson 1984; and Digital archive). The excavation methods of Armstrong within the cave have recovered large quantities of micro-faunal remains including species such as bats, which are of a similar size. In the circumstance, it seems unlikely that the data is biased by the non-recovery of remains of these species and in such circumstances the surviving material is likely to reflect the actual distribution within the sediments.
A total of 394 vertebrate bones have been identified as either Common Frog or Toad (Figure 104 and Digital archive). Remains were also recovered during excavations by the author. During this work an area used by Armstrong for storage of remains and limestone clasts was cleared; Figure 5) The sediment from this area was sieved and hand sorted to recover micro-vertebrate remains. A total of 4293 amphibian bones were recovered (see Digital archive), which unfortunately do not have a stratigraphic context. Additionally, excavations by the author within the rear section recovered amphibian remains from calcrete blocks within the 'inner chamber' and sediments from the rear infilling section. These have yet to be fully identified. The main point of referencing these remains is that it provides comparative information relevant to Armstrong's excavation. There is no doubt that his very methodical technique recorded huge quantities of micro-faunal remains. In this instance, his recovery and annotation of 394 amphibian bones (above) does not compare favourably with the 4293 recovered by the author from the stored 'spoil' sediments.
Analysis of the Frog bones from Armstrong's excavations shows that limb parts are the most frequent but are associated with axial and cranial parts. The species occur in very low numbers between stratigraphic levels 7 to 14 and increase dramatically within level 2. Most of the surviving bones are complete and have little sign of damage by predators or fracturing within the sediment.
Their stratigraphic distribution differs from that known for predators such as raptors, Red Fox and Wolf, which suggests residence by the species. The single exception is seen within level 2 where the significant increase in number, mainly limb parts, may be due to raptor predation and where species of both Falcons and Owls occur. It is very likely that the species was both resident and also the prey of small carnivores. The difficulties of access to this area of the cave make it extremely unlikely that the remains are the result of Raptor and Owl predation.
A total of 8 species of fish have been excavated from the cave and their distribution occurrence is shown in Figure 105. A total of 1338 fragments are described, 1029 of which are scales. Much of the identification was undertaken by Dr Mike Wilkinson (Sheffield University) and represent 26 MNI. The excavated material includes 34 Pisces bones and scales that have not been identified. The group is mixed in the sense that they require differing habitats. Eel (Anguilla anguilla) and Salmon (Salmo sp) inhabit clean rivers while Flounder (Platichthys flesus) prefers estuarine conditions and each species requires access to the sea.
Grayling (Thymallus thymallus) and Pike (Esox lucius) inhabit a variety of water condition in rivers and lakes. Both Chub (Leuciscus cephalus) and Carp (Cyprinid sp) are found in similar habitats and are of European origin (Maitland 1977). Their presence within the cave indicates that both fast and slow flowing rivers, some with access to the sea, occur both locally and regionally.
Grayling is known from 1523 bone and scale fragments. A total of 332 have sufficient dimensional detail to be incorporated within the reconstruction (21% of the total) and a further 78% of the remains can be located to the 'passage levels'.
Grayling remains are concentrated within the area 58-70ft (17.6-21.3m) north from the entrance and between 6-12ft (1.8-3.6m). Their distribution indicated that they are within the mound of sediment underneath the roof rupture. At the 9ft (2.7m) depth they seem to spill into the 'inner chamber', which in this area and at this level probably contains Quaternary deposits. Remains are dominated by scales that in several instances, especially in the lower levels within the rear of the cave, seem to belong to the same individual.
The species are found in virtually all stratigraphic levels. In levels 8 to 13, Grayling, Eel and Pike are the most frequent. Levels 3 to 7 see the appearance of low numbers of the additional species Chub, Perch and Flounder.
Figure 105: Fish species (Pisces) temporal distribution (source data: Fish, Grayling, Pike)
Figure 106: Fish species, body part selection (source data: Digital archive)
The preservation of body parts is very distinct and is shown in Figure 106. Scales are by far the largest number of remains, although these are likely to belong to a few individuals. Low numbers of cranial and, more rarely, axial parts are preserved. The relatively high numbers of scales within the lower sequence are associated with at least one, large, European Brown Bear. The removal of scales and the virtual destruction of internal bone are characteristic of bear predation where large parts of the carcass may have been swallowed, unchewed, and bony parts passed in faeces. This style of survival is very distinct within stratigraphic levels 8-12.
Figure 107: Pike (Esox lucius), well-preserved mandible and maxillary fragments from stratigraphic level 12, Pin Hole Cave (Image courtesy of Manchester Museum)
Direct evidence of predation is evident on at least one maxillary fragment of Pike (Figure 107). These associated fragments, from stratigraphic level 12, probably belong to the same individual. The maxillary fragment has evidence of two medially located bite marks, which from their form appear to be those of a small carnivore canine. There is no direct evidence that would indicate human interest in any of the species. Given their location, it is also unlikely that they indicate Falcon or Eagle predation. The species Pike and Grayling both have distributions that are within the cave rear and are associated with levels that contain Middle Palaeolithic lithic assemblages and the remains of European Brown Bear, but there is simply not sufficient distribution detail to identify the predator.
6.13 Interpretation and summary - the vertebrate populations
The vertebrates described represent a Quaternary faunal population of immense richness and diversity and which colonised, adapted and survived great variation in environmental conditions throughout an immense period of time. The evidence is very unusual in that it contains many species that are not commonly found within similar sites within the UK. These species also illustrated a range of habitat preferences that are of considerable interest for studies of biome and ecotone changes within the Quaternary.
The variation in genera and species has been considered with particular reference to a wide range of taphonomic circumstances that help explain how such a huge quantity of vertebrate remains have partly survived within a relatively small and narrow cave.
Interpretation and summarising taphonomic events within the cave is difficult. The context, already discussed, involves 24 cubic metres of void within the cave, semi-filled with sediment and biological remains. The density of the latter is in the region of 22,516 vertebrate bones and fragments, which average an occurrence of 938 per cubic metre. The issue is slightly more complex as analysis of the remains suggest that for the most part the excavated finds represent 10% or less of the total activity. Many of these live animals have lived, moved and died within this small environment, actions that would have had an effect upon the cave interior and infilling deposits. In the circumstances, it is clear that the cave interior is an area of extensive bioturbation, little of which was recorded by excavation and this has developed within a background of a slow and gradual weathering and movement of decaying limestone and introduced sediment. The sheer density of remains suggests that all aspects of the evidence have been disturbed by this biological process to varying degrees. Despite the complexity, the analysis of the vertebrate population and the evidence of taphonomic effects on the remains suggest the appearance and development of a new ecosystem by ungulate behaviour is a causal factor in Neanderthal appearance in the hills of Eastern England. This theme is considered more fully in later discussion of an ecological niche (Section 10).
It has long been realised that Hyaena predation and feeding practice has been a significant taphonomic factor within Pin Hole Cave's history. Direct evidence of breeding in the form of a foetal skeletons considerably adds to this picture. This study highlights the role of several other carnivores that have left a taphonomic imprint within the vertebrate record. The circumstances of European Wolf predation, European Brown Bear residence and the activities of Red Fox have been significant.
Analysis of Perissodactyla remains has demonstrated the importance of Woolly Rhinoceros as prey of Hyaena and to some extant Neanderthal groups. The abundance of Reindeer and its association with humans and carnivores clearly demonstrates that it is an important food resource as well as a major source for raw materials.
The variations in the components of faunal populations are of considerable interest, and contrast with evidence from other Quaternary sites.
Use the navigation bar (top) to go to other sections of this publication
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Internet Archaeology content is preserved for the long term with the Archaeology Data Service (ROR). Help sustain and support open access publication by donating to our Open Access Archaeology Fund.
 Home Summary
Home Summary
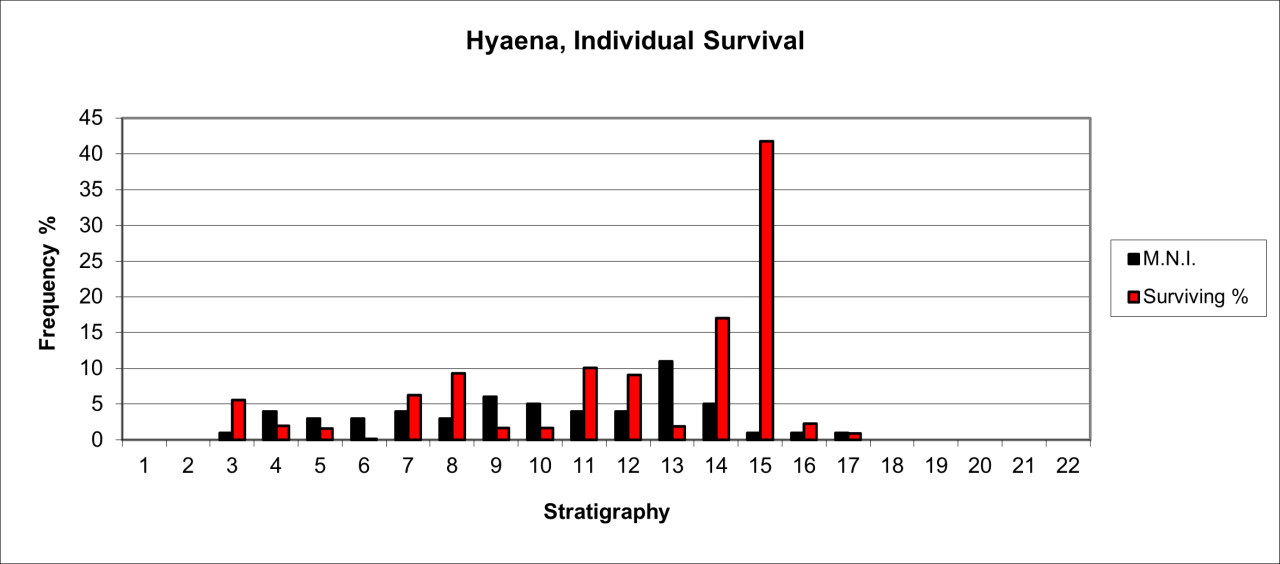


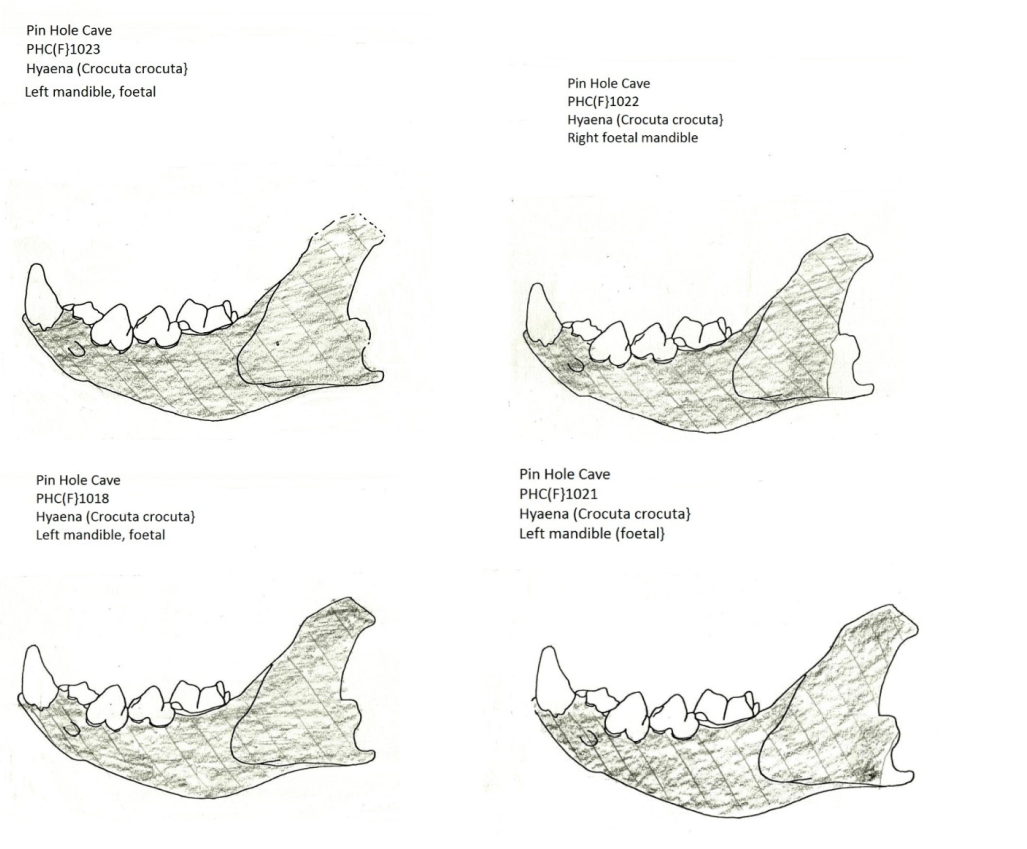
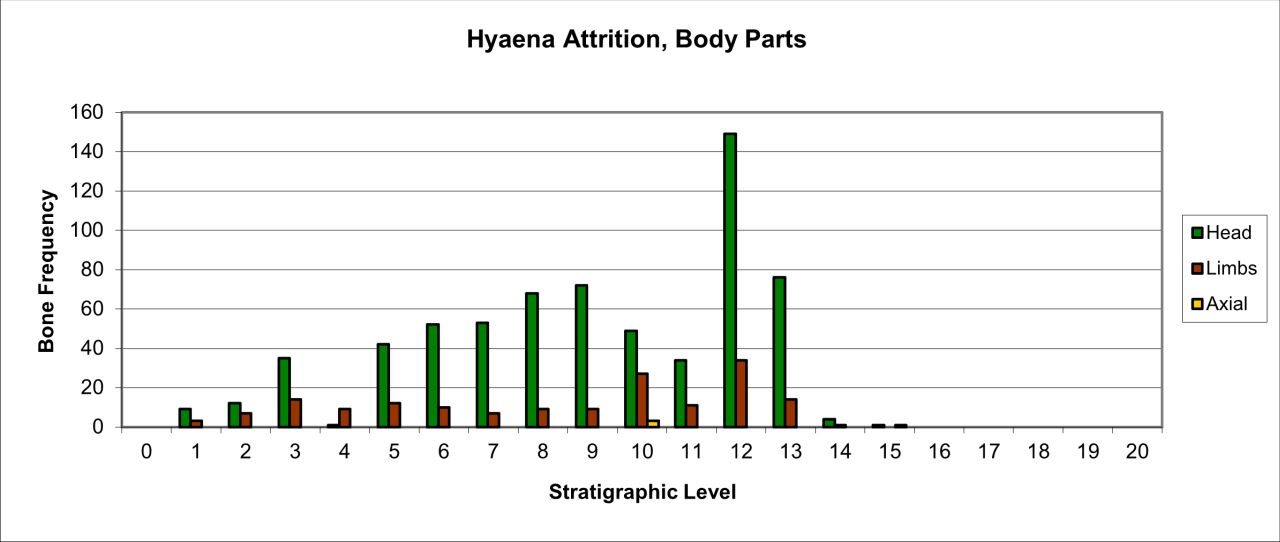
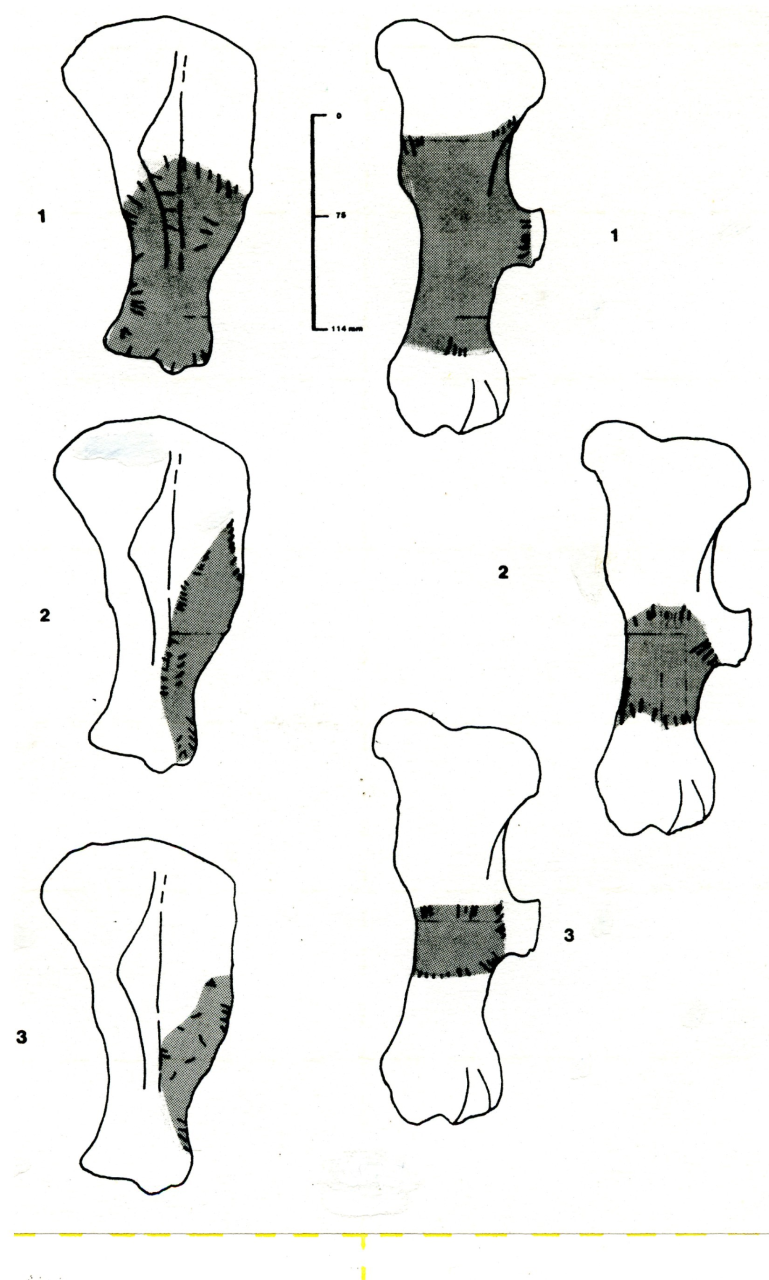
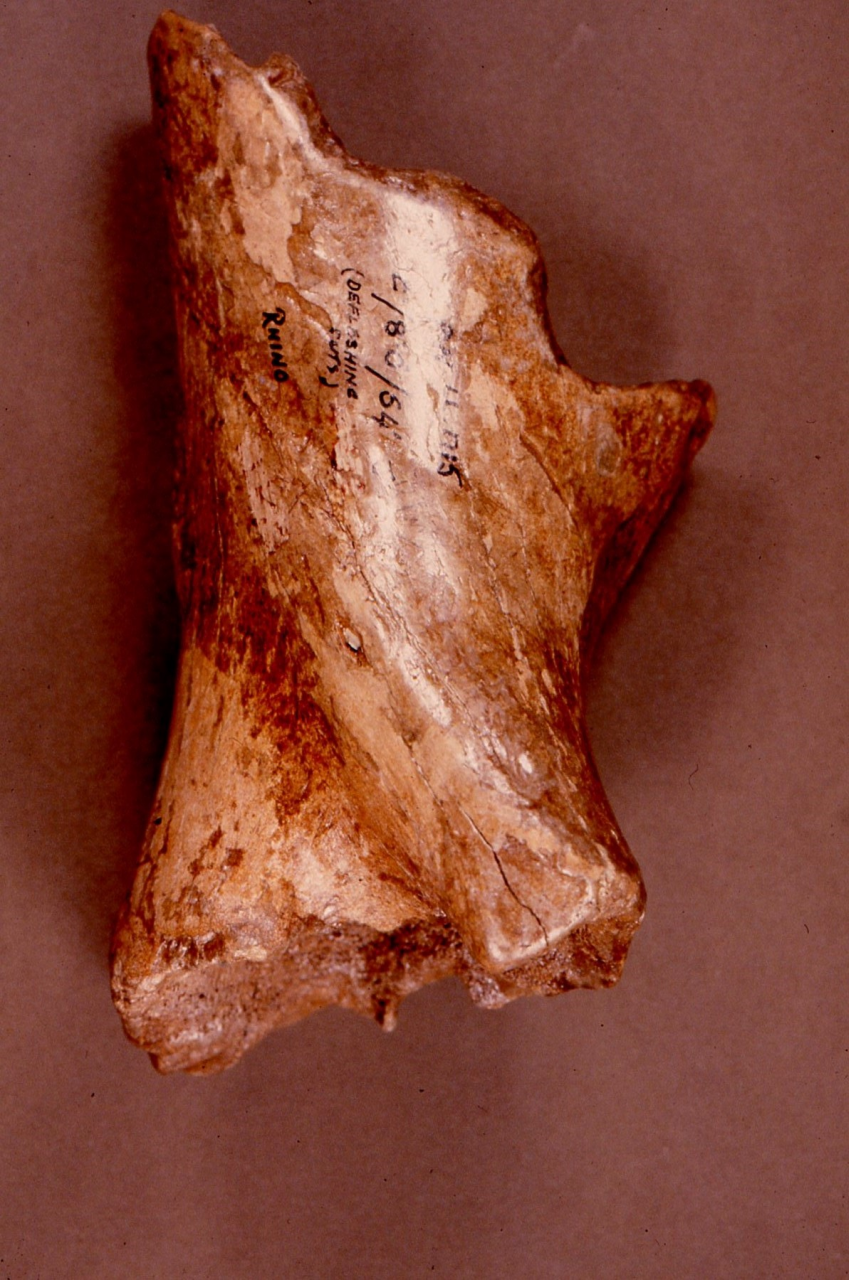
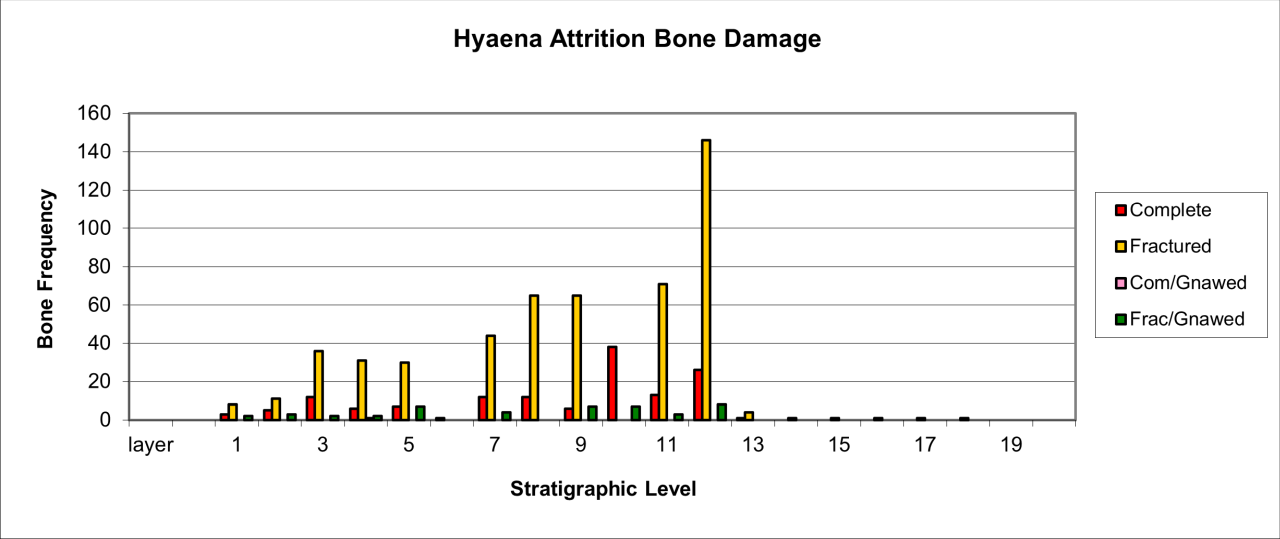

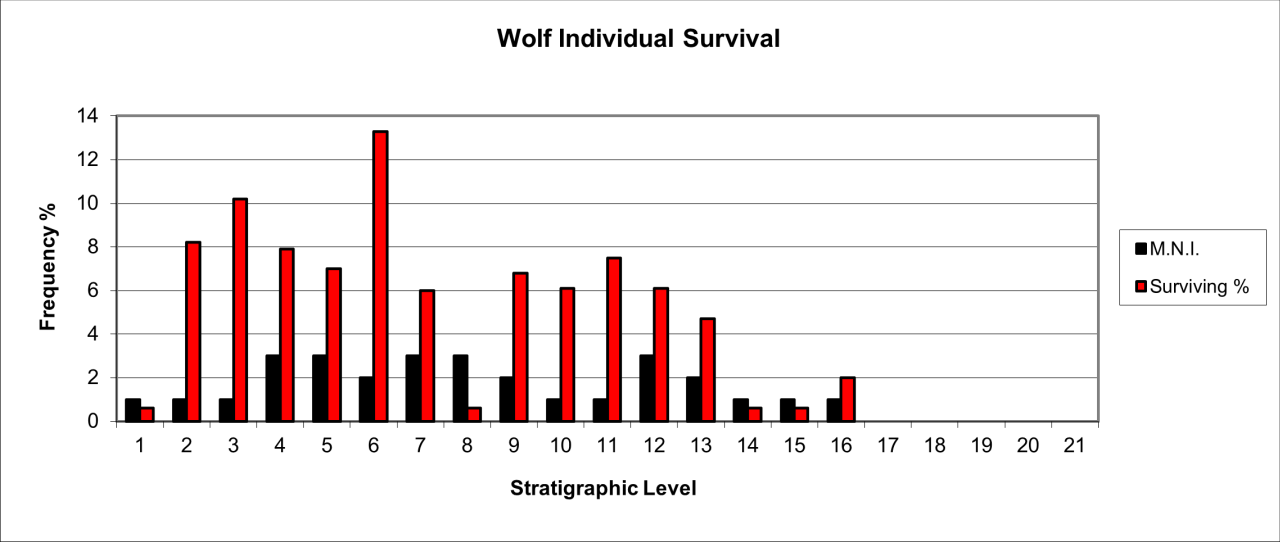
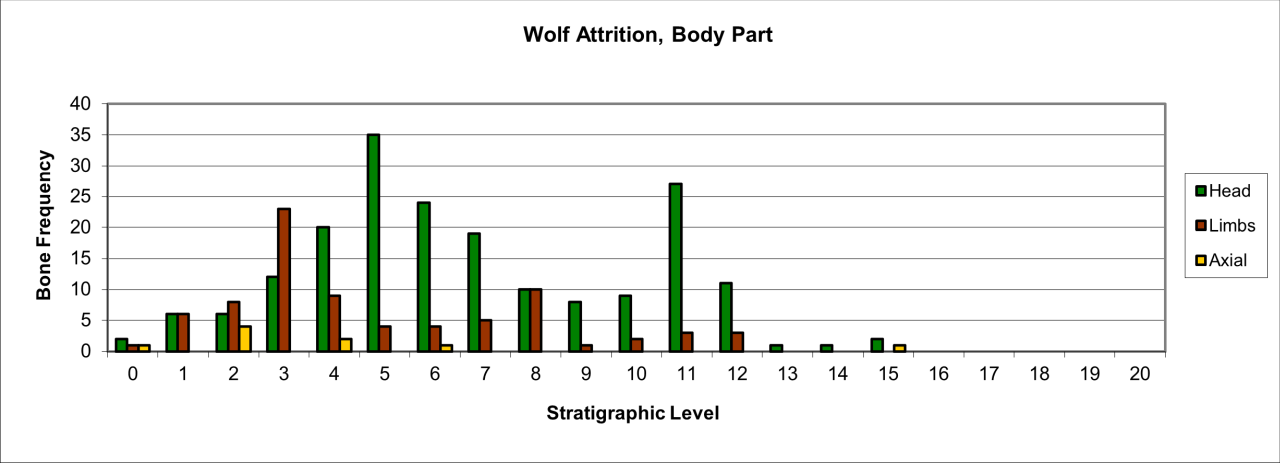
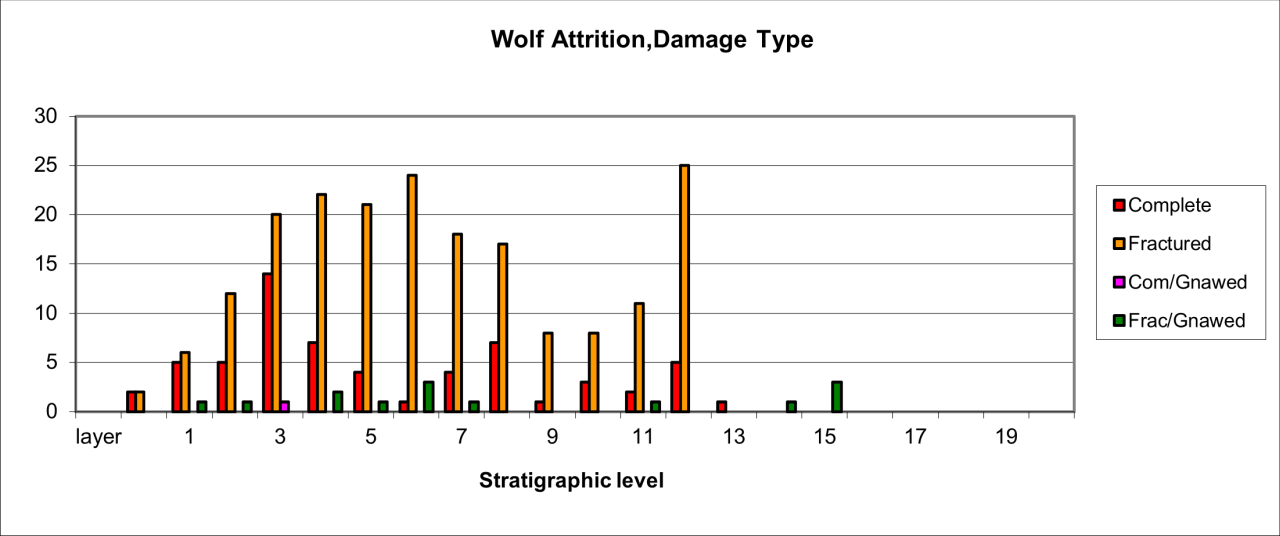
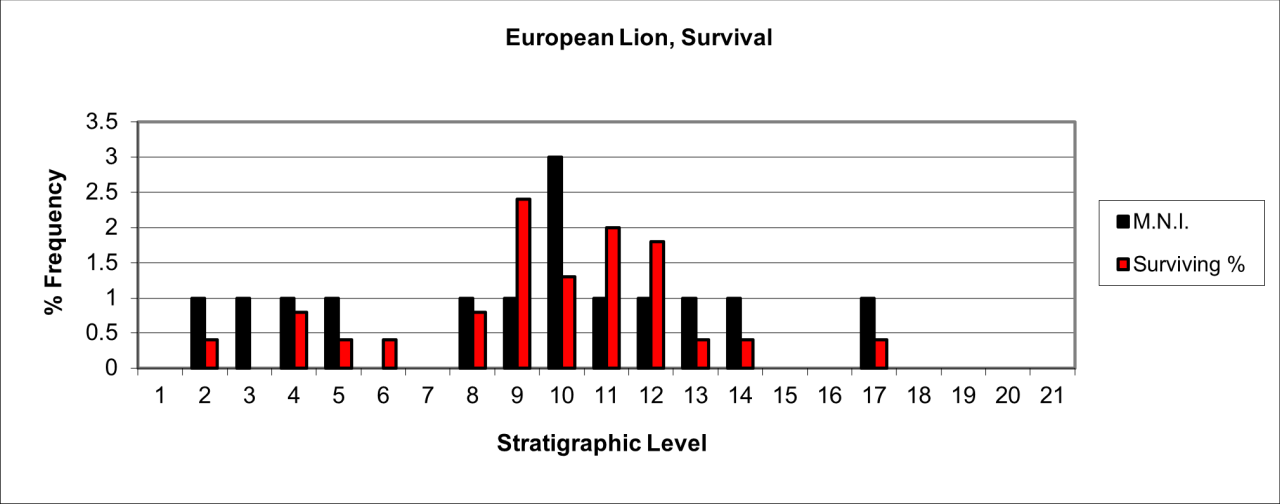
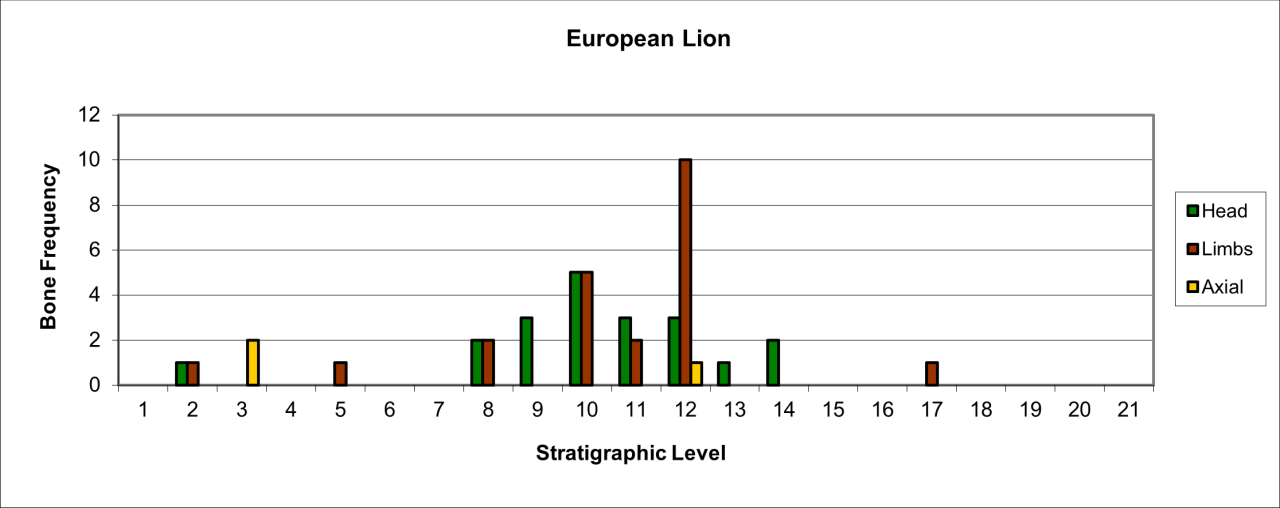
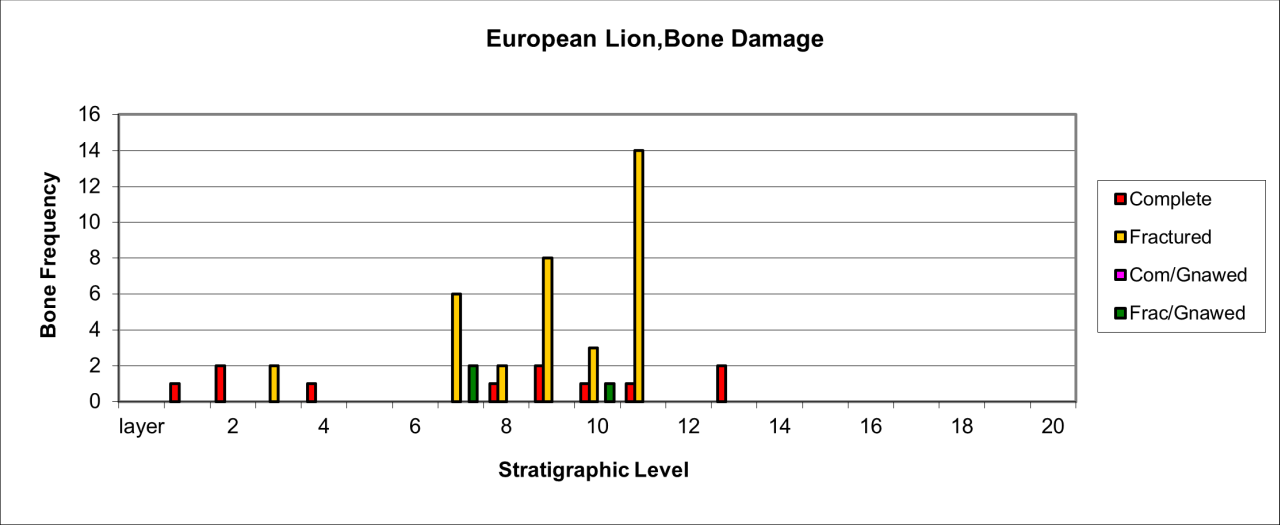
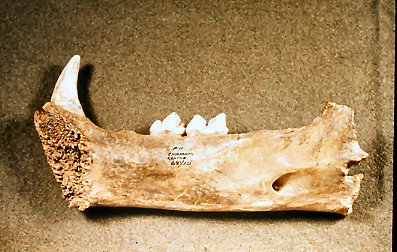
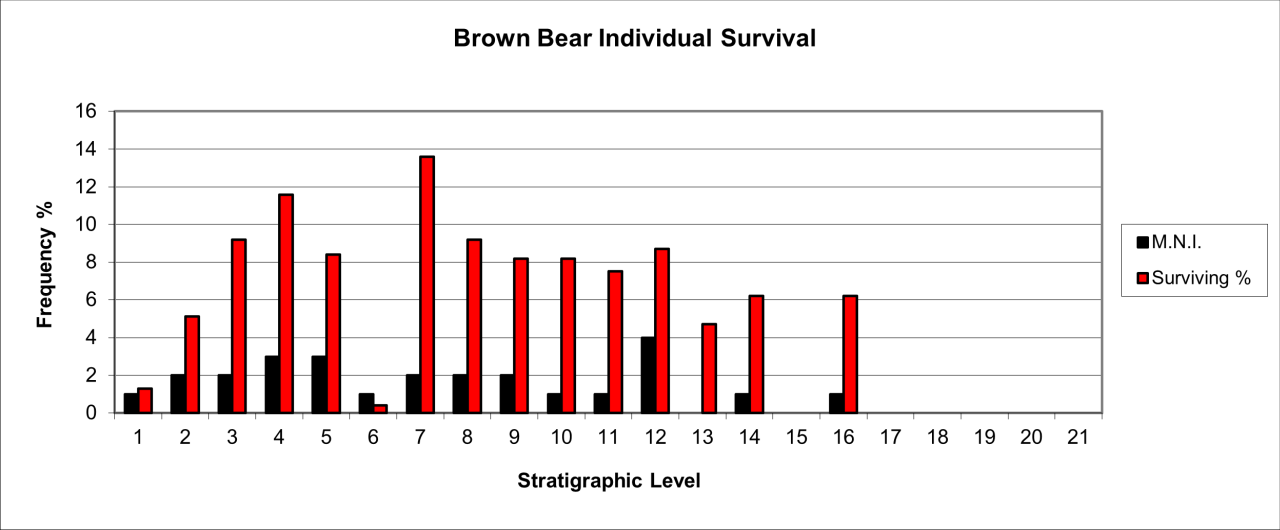
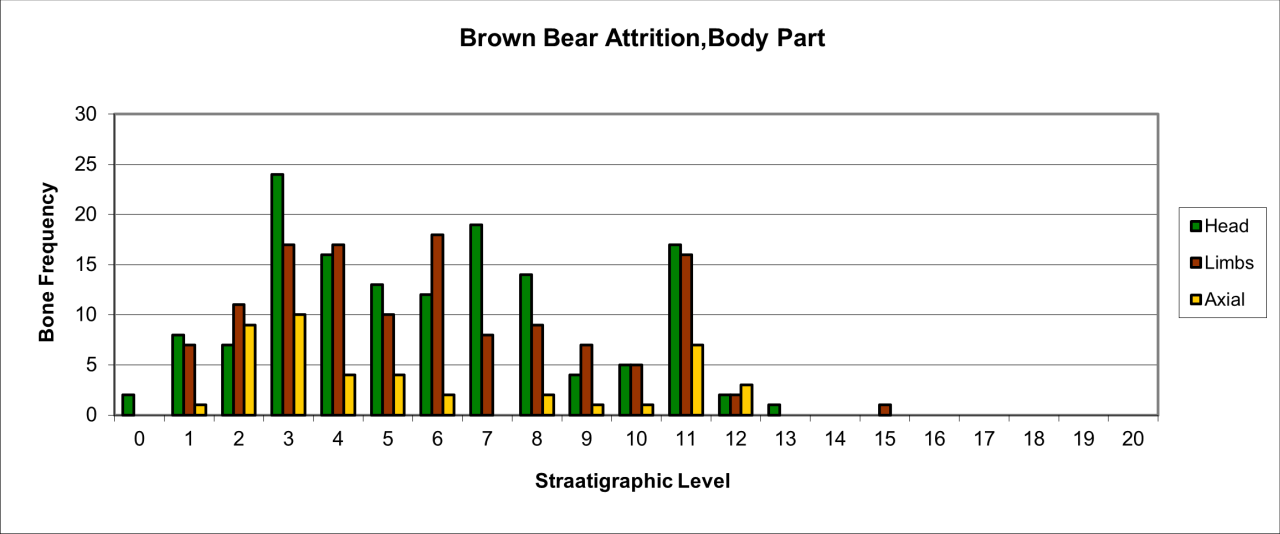
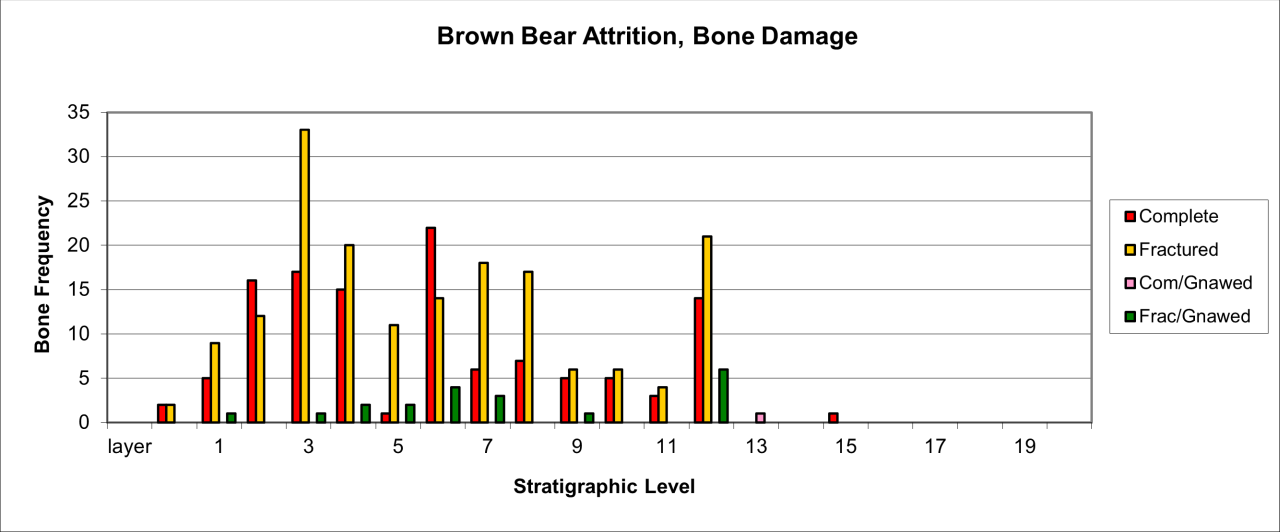
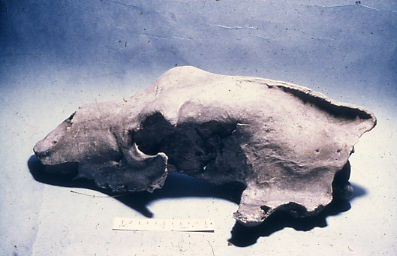
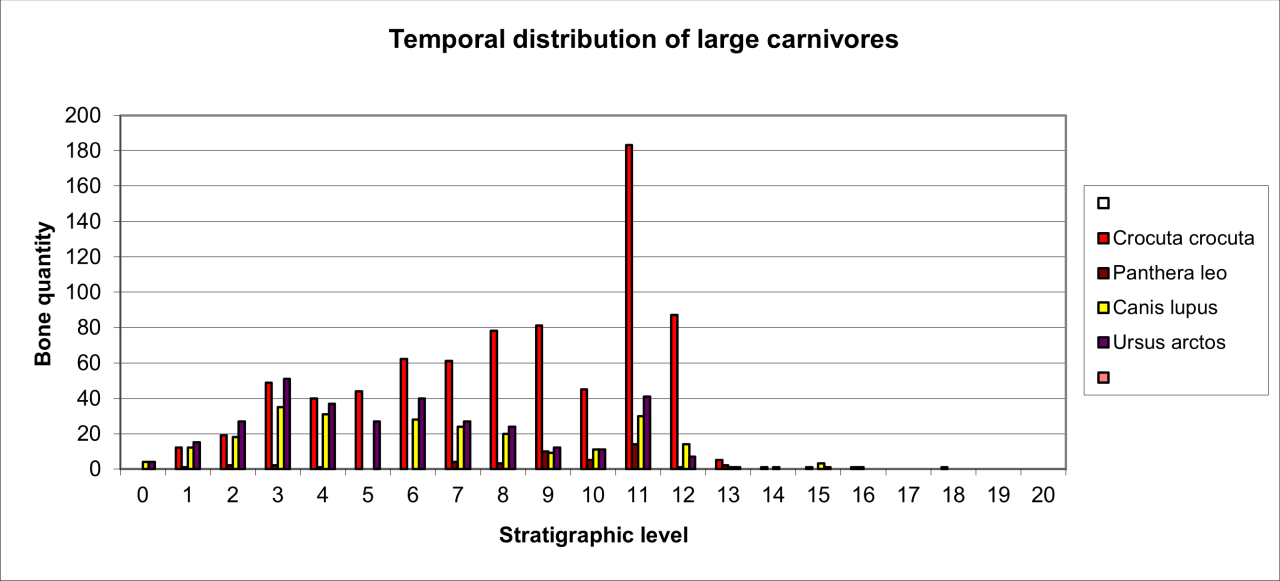
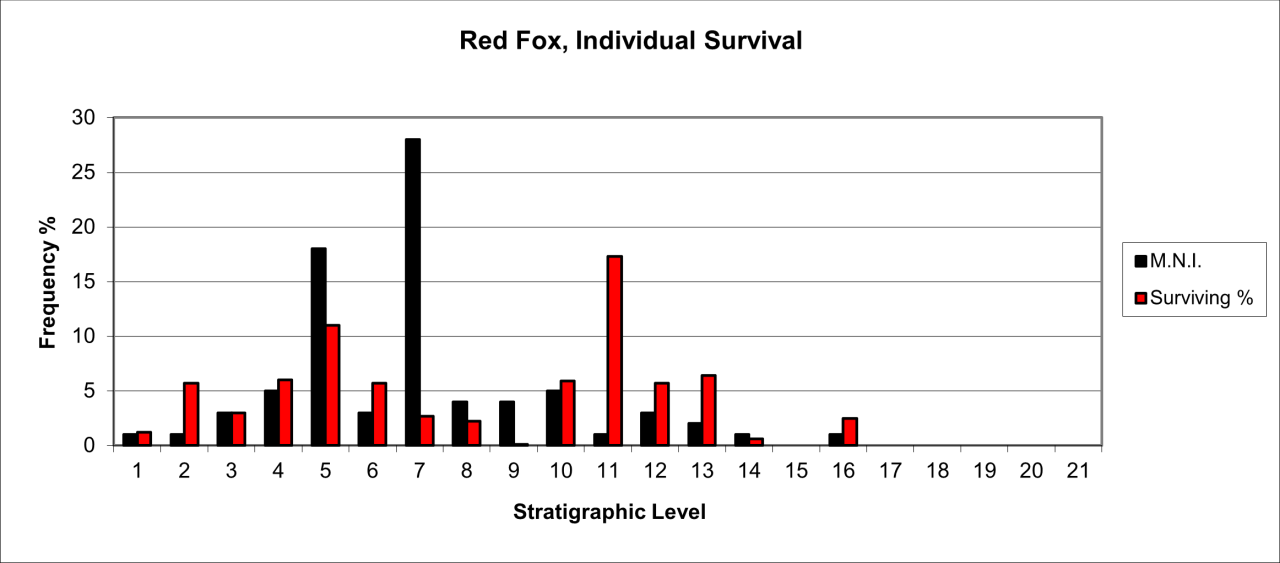
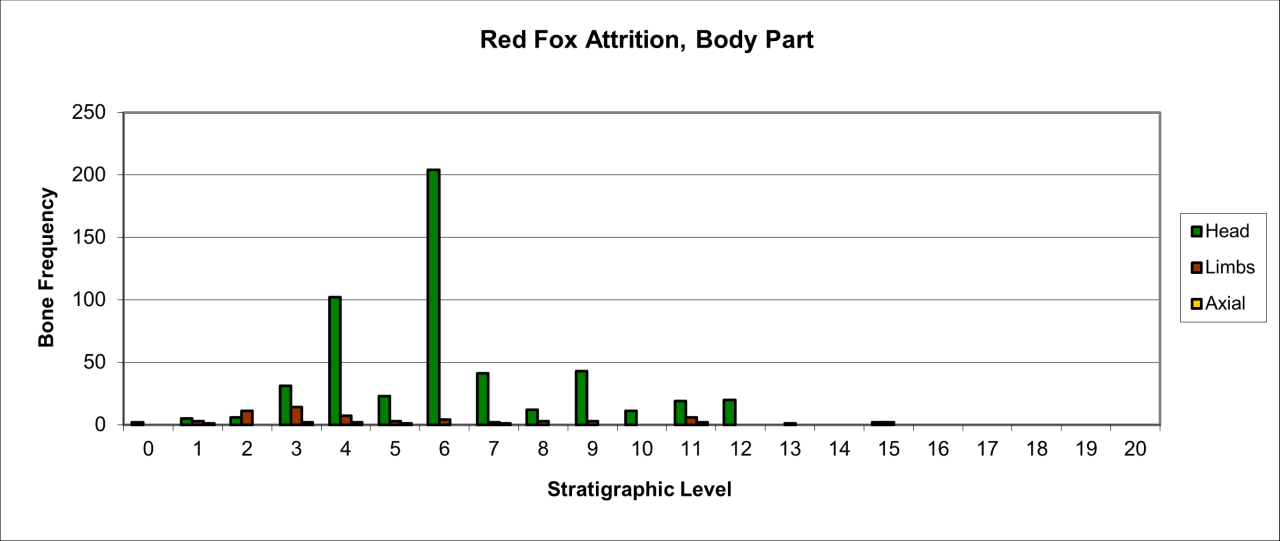
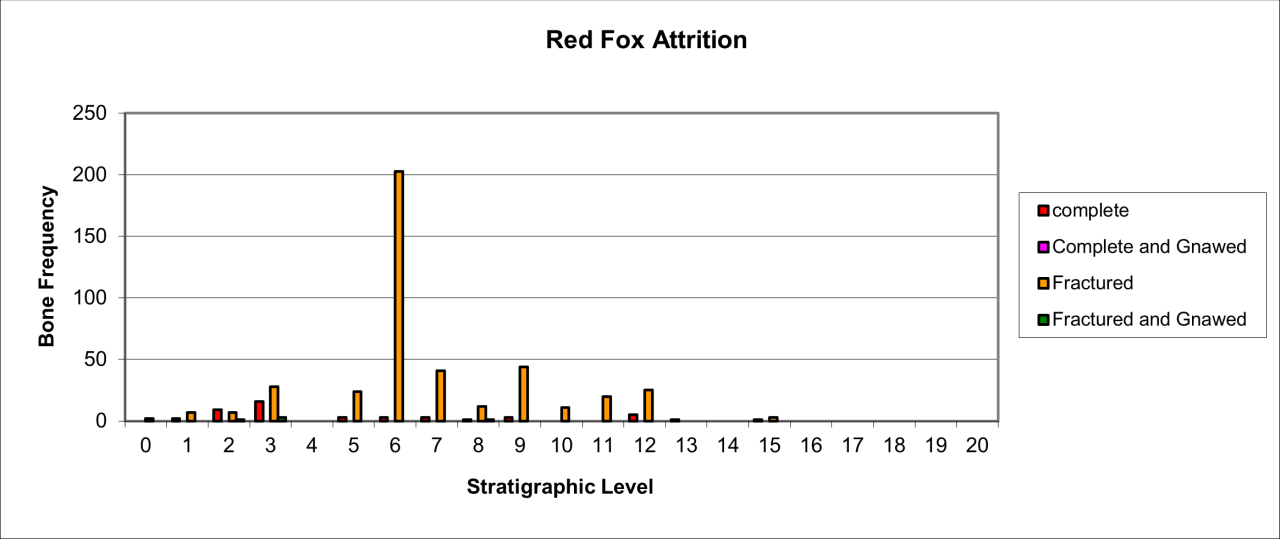
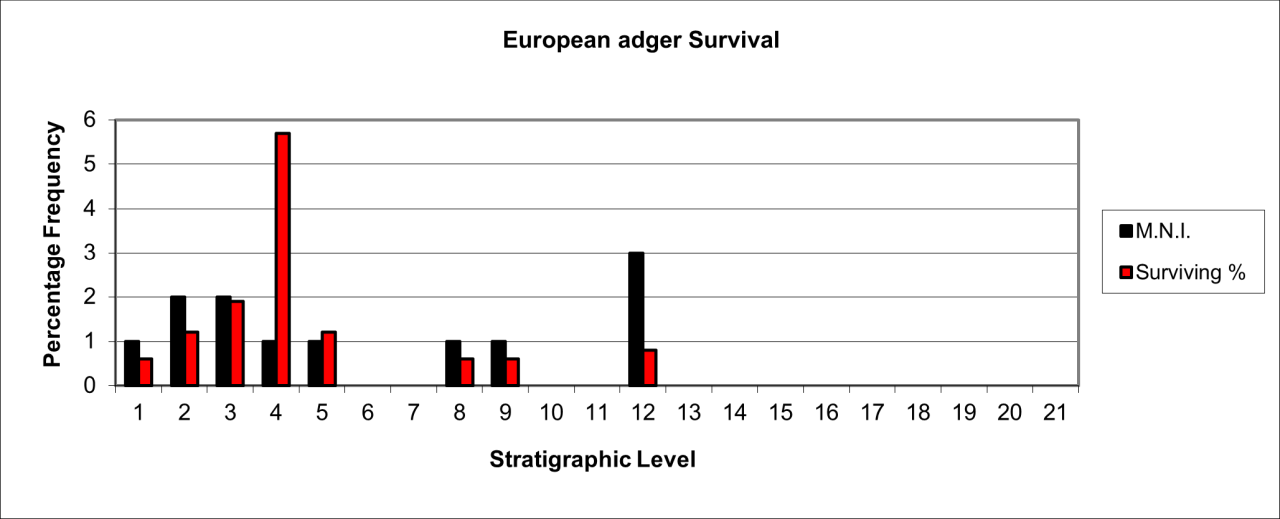
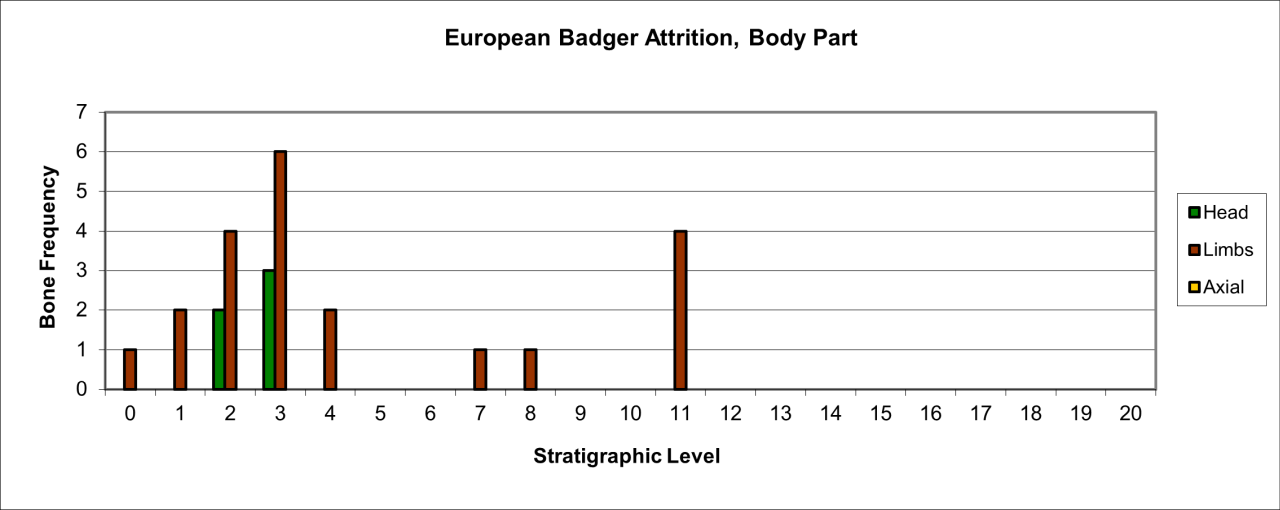
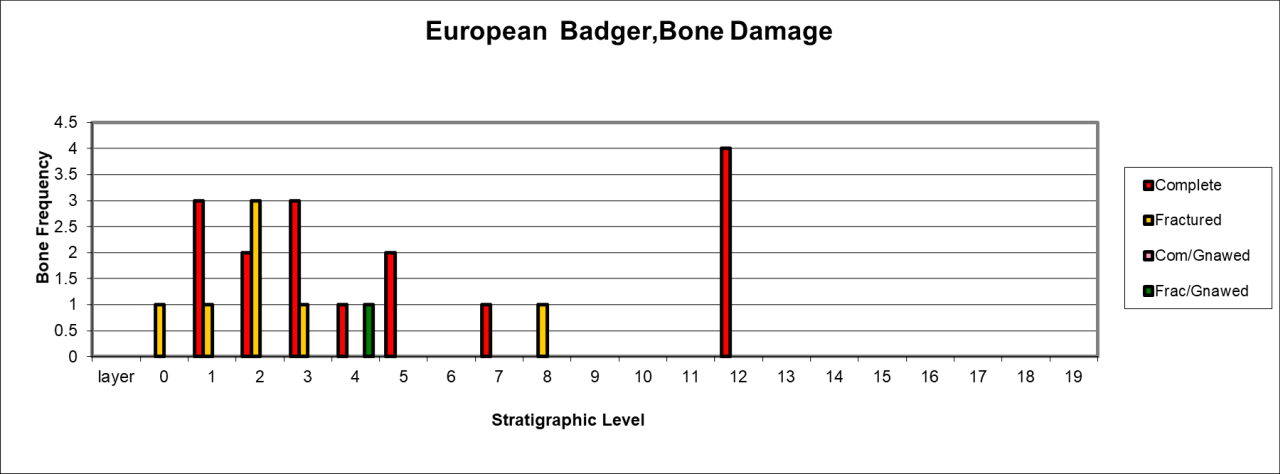
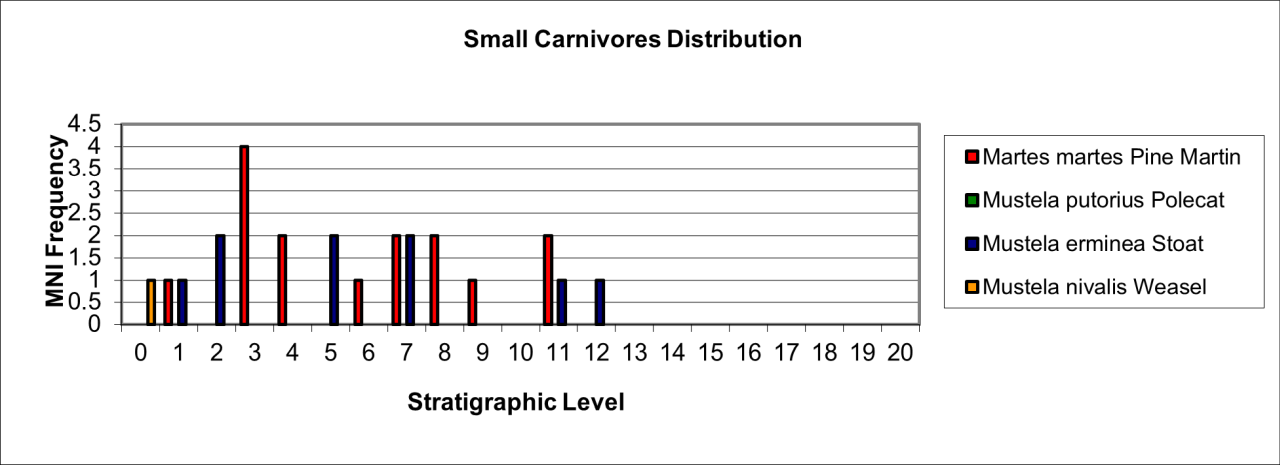
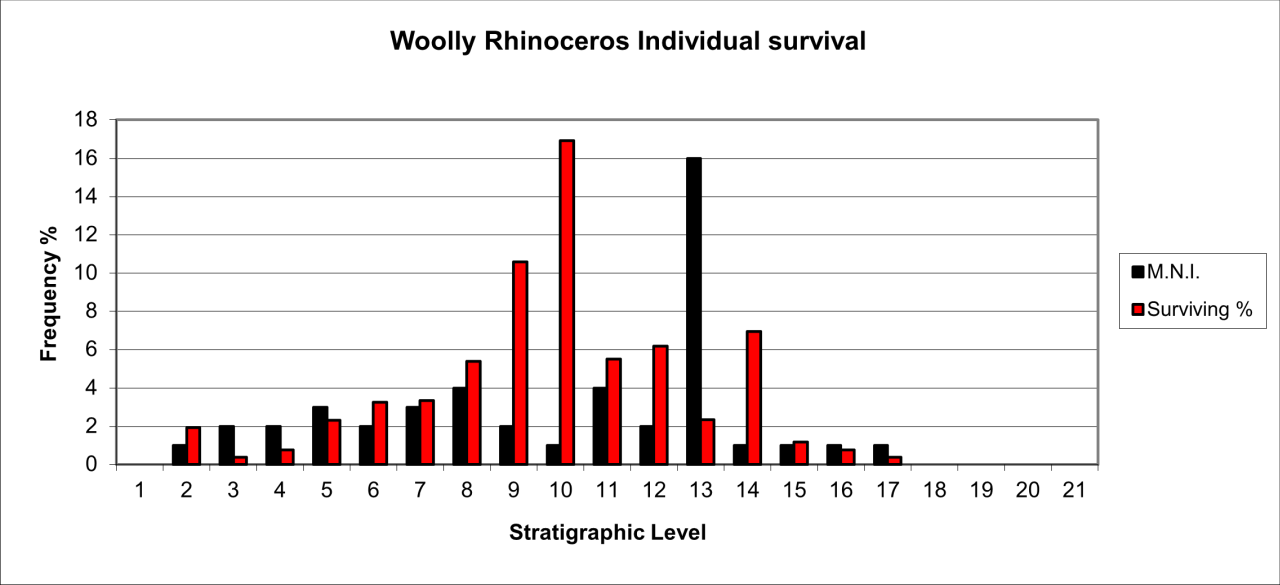
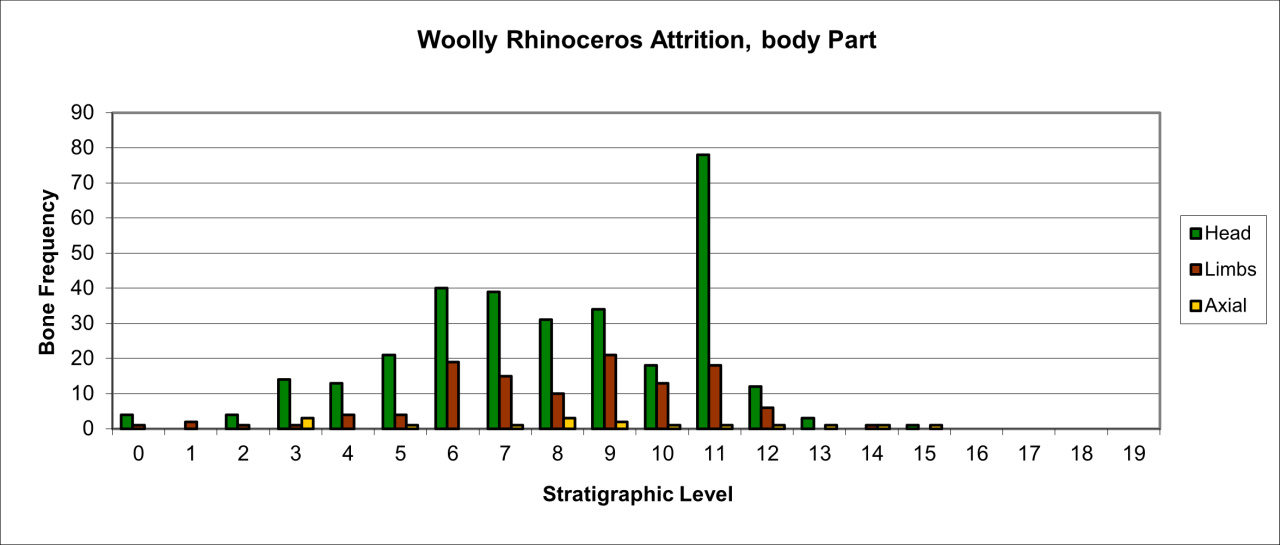

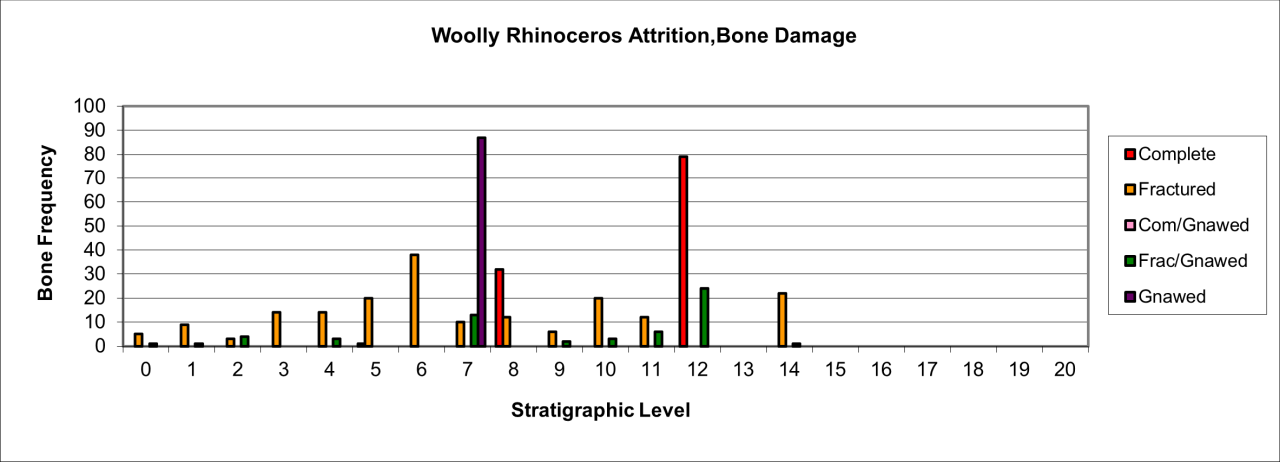
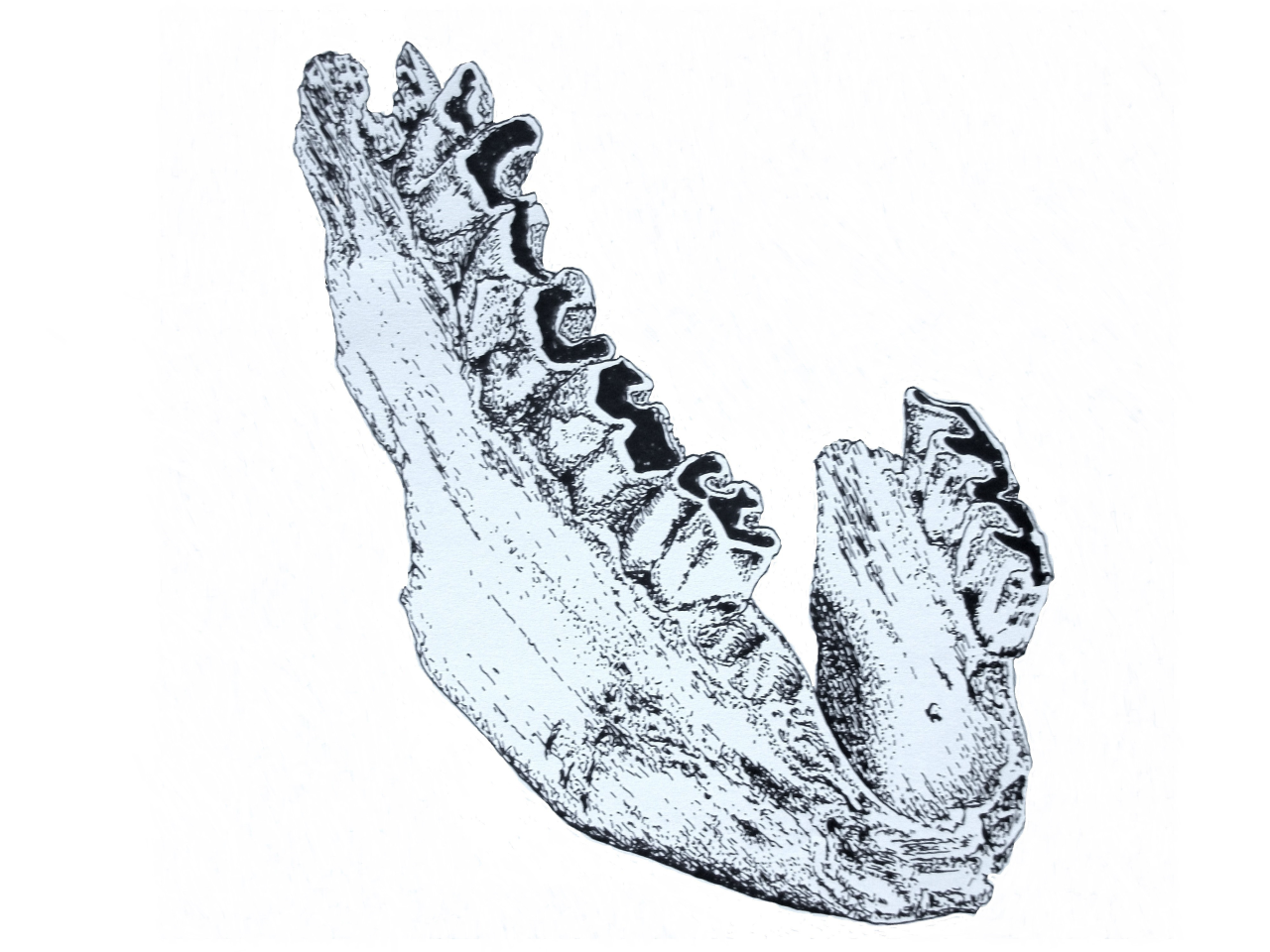


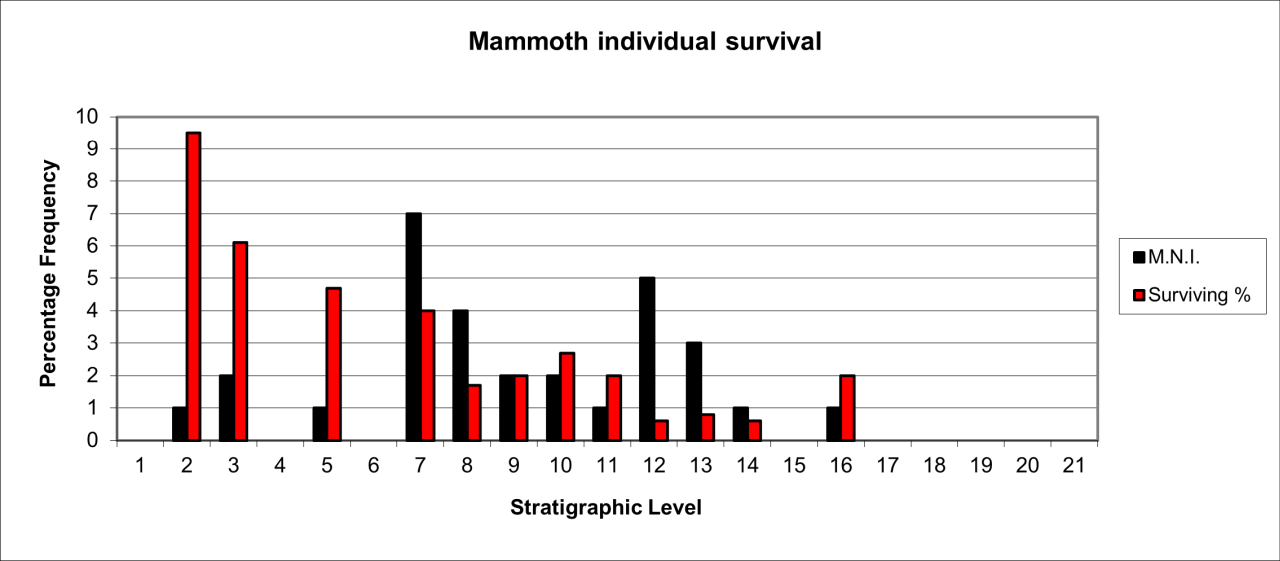
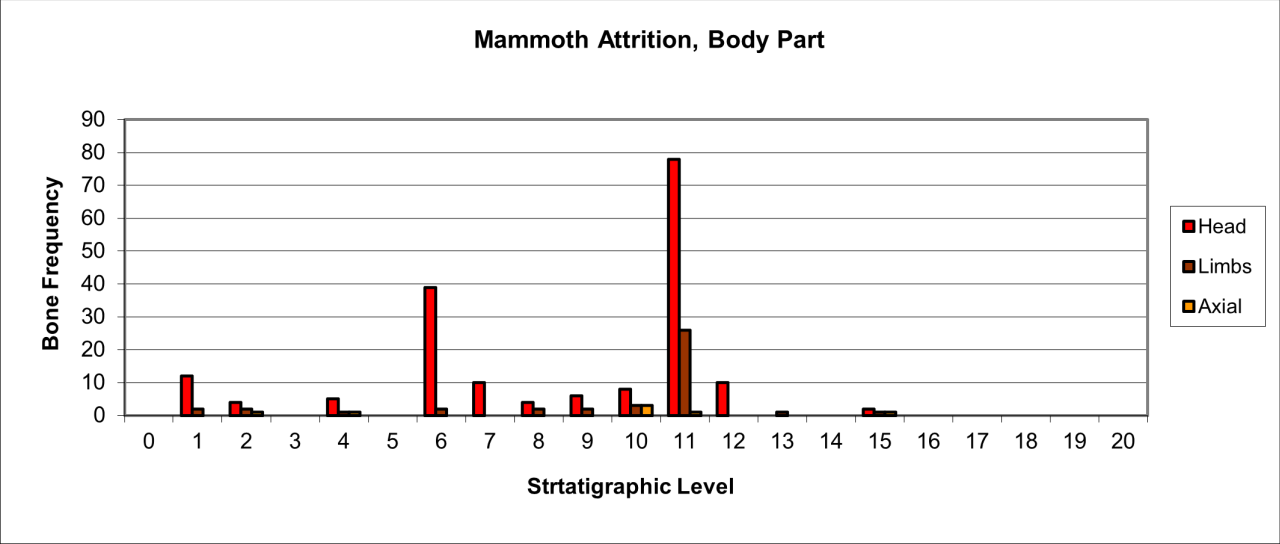
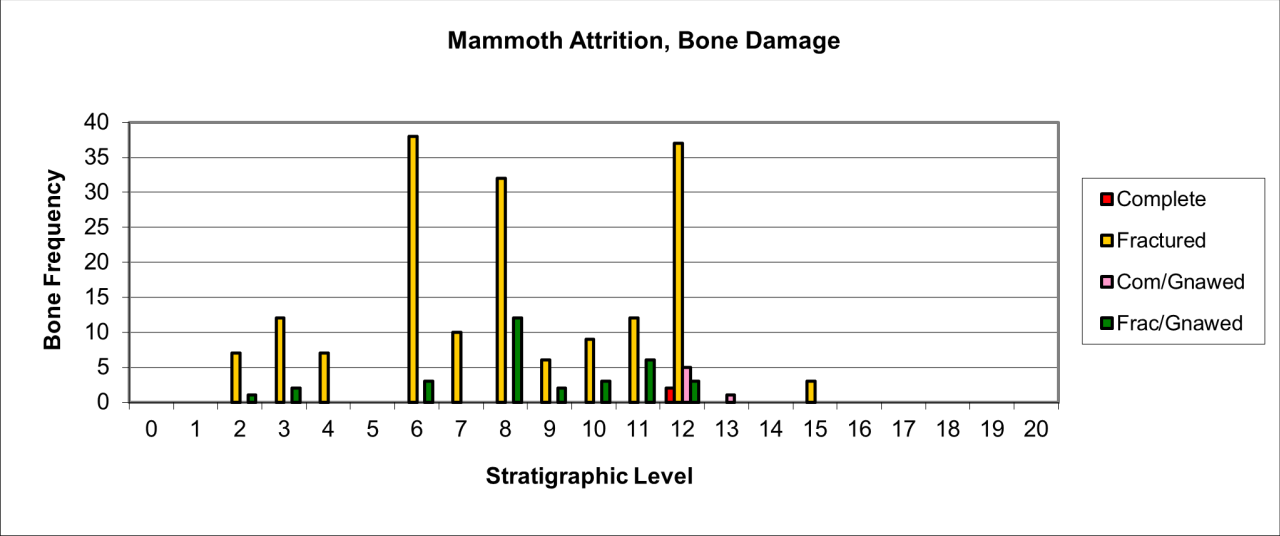

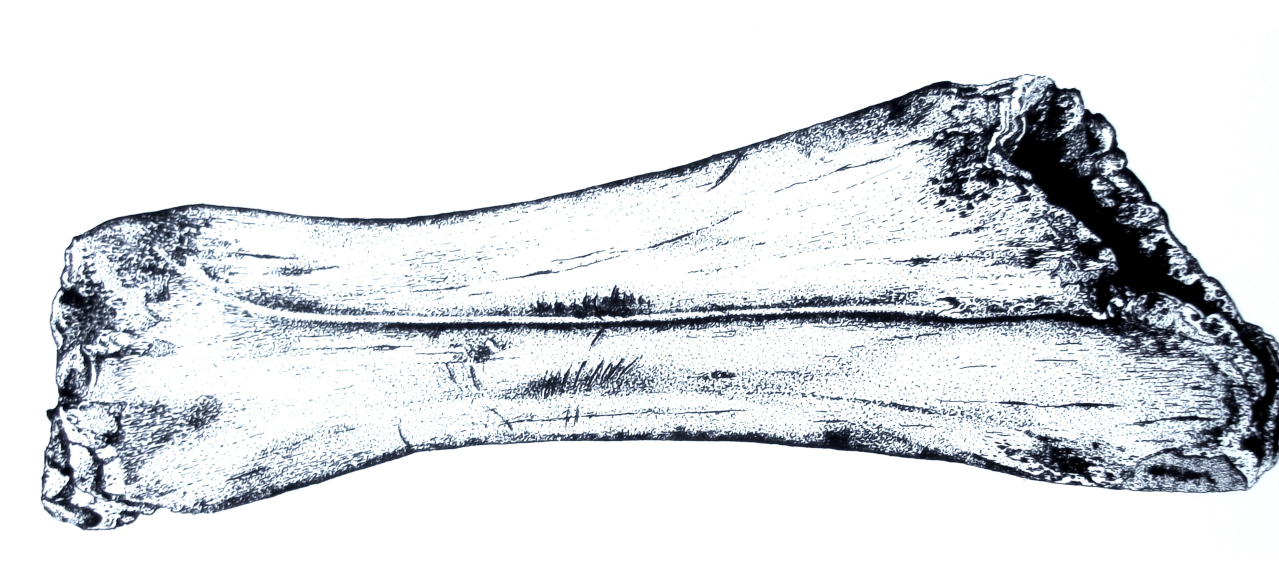


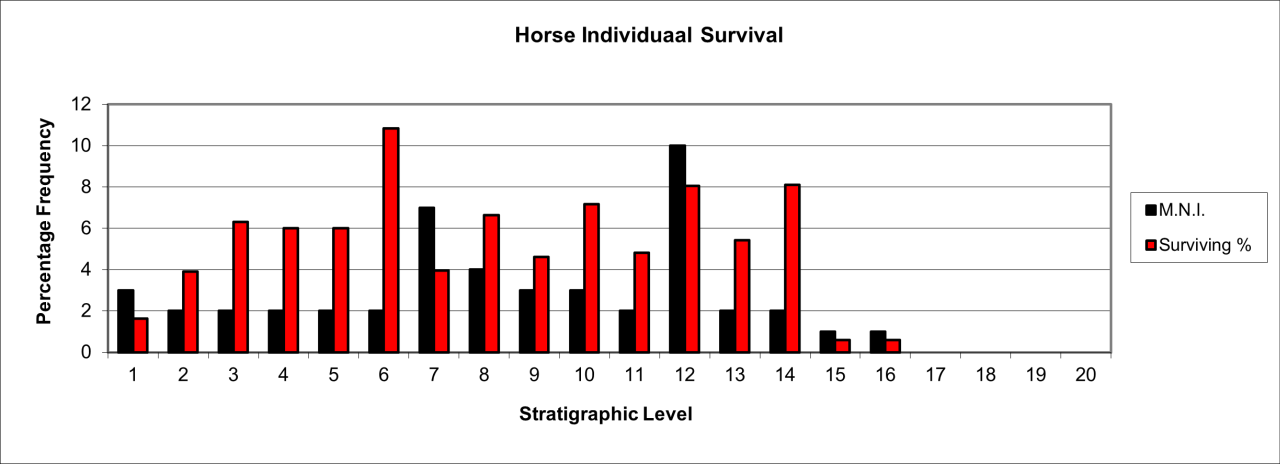
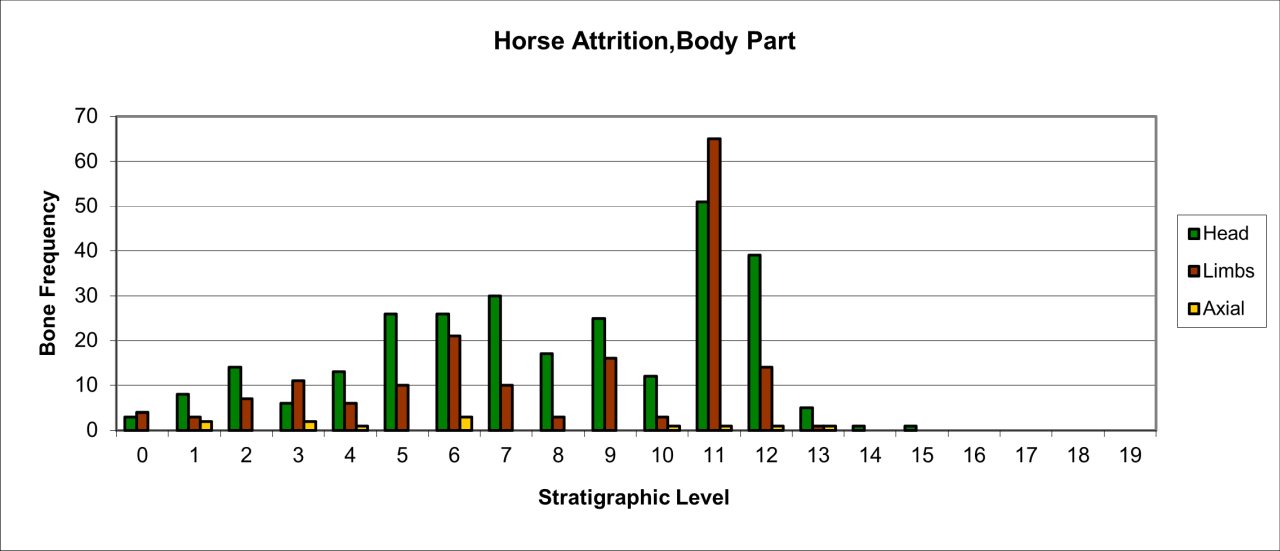
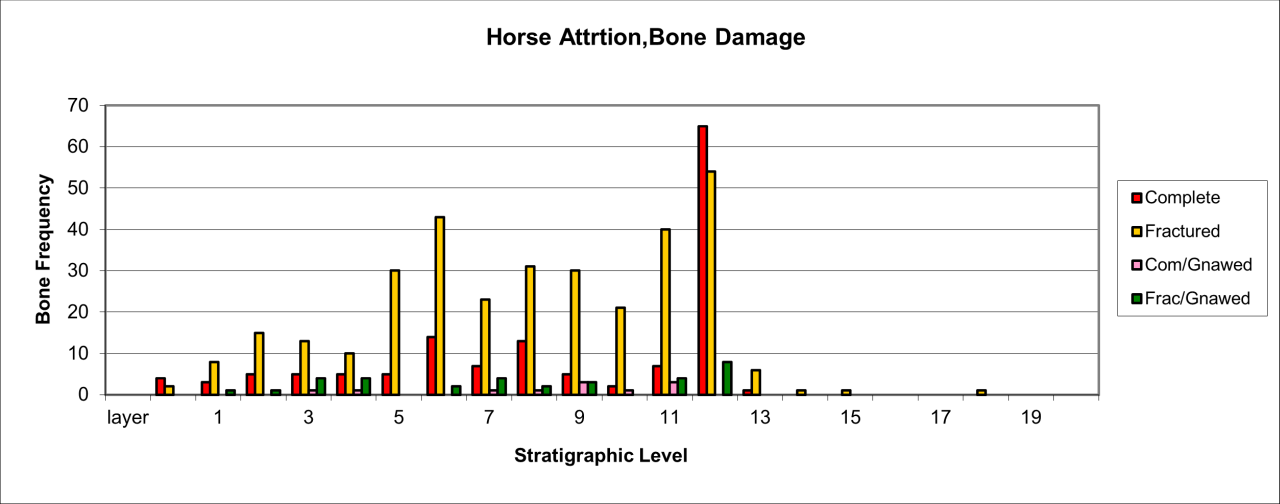
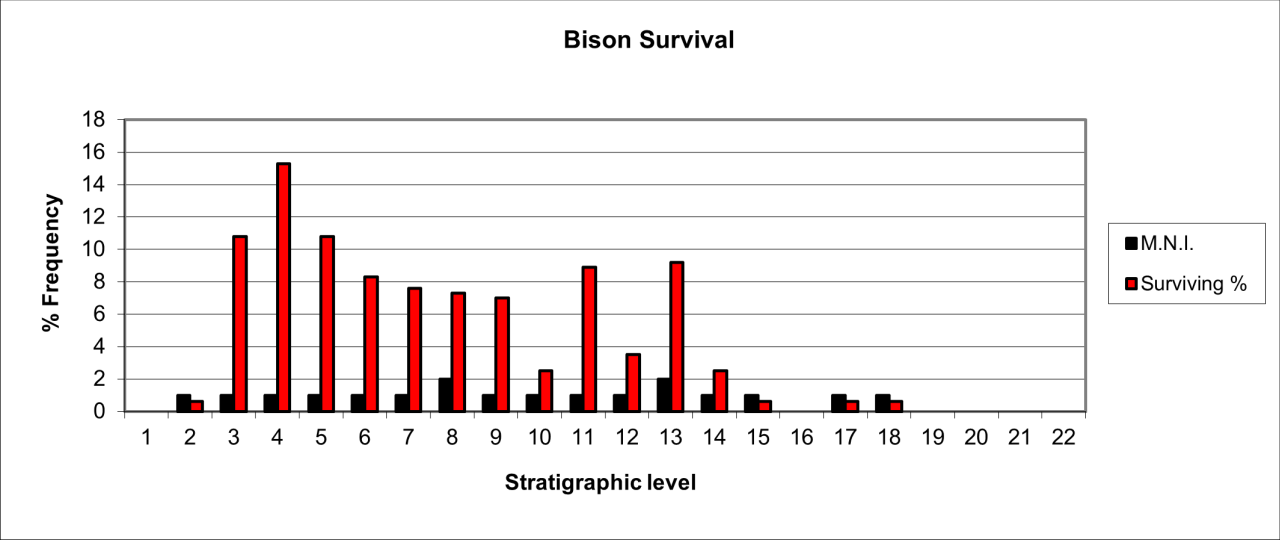
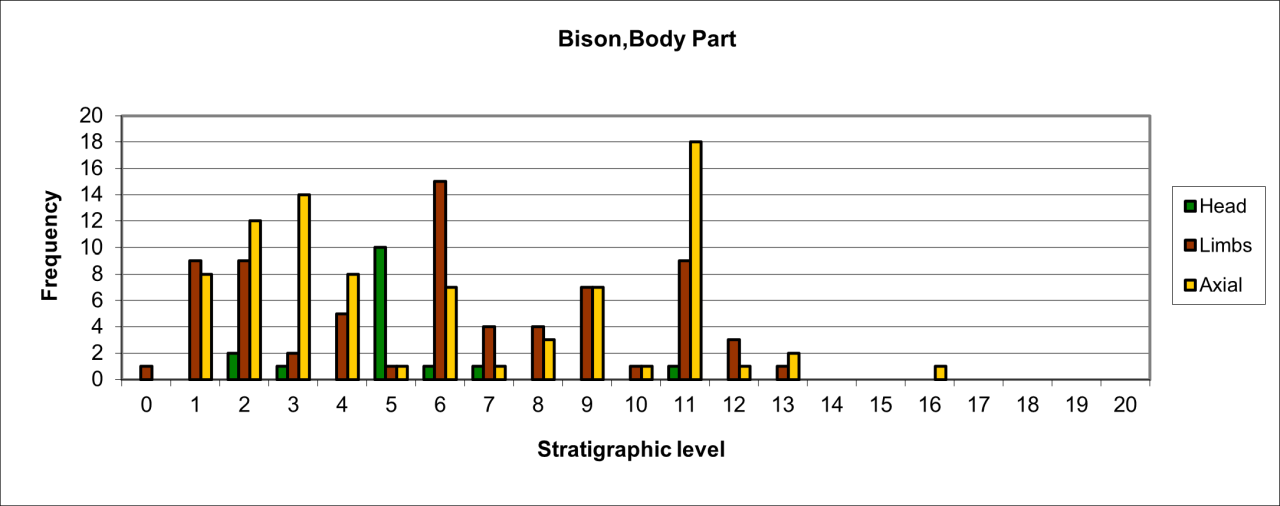
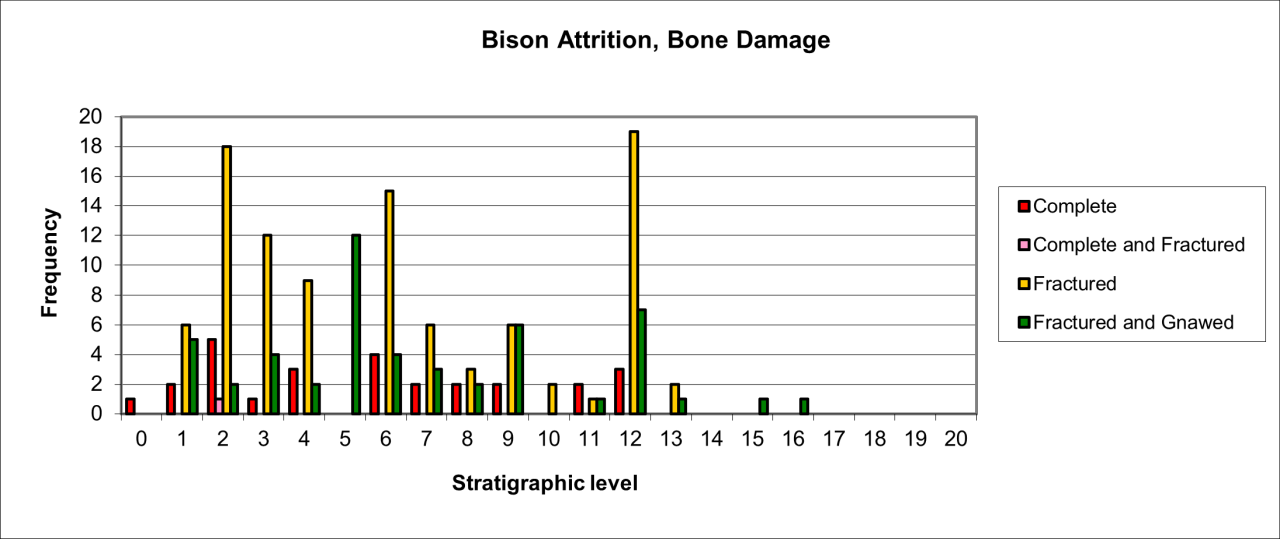

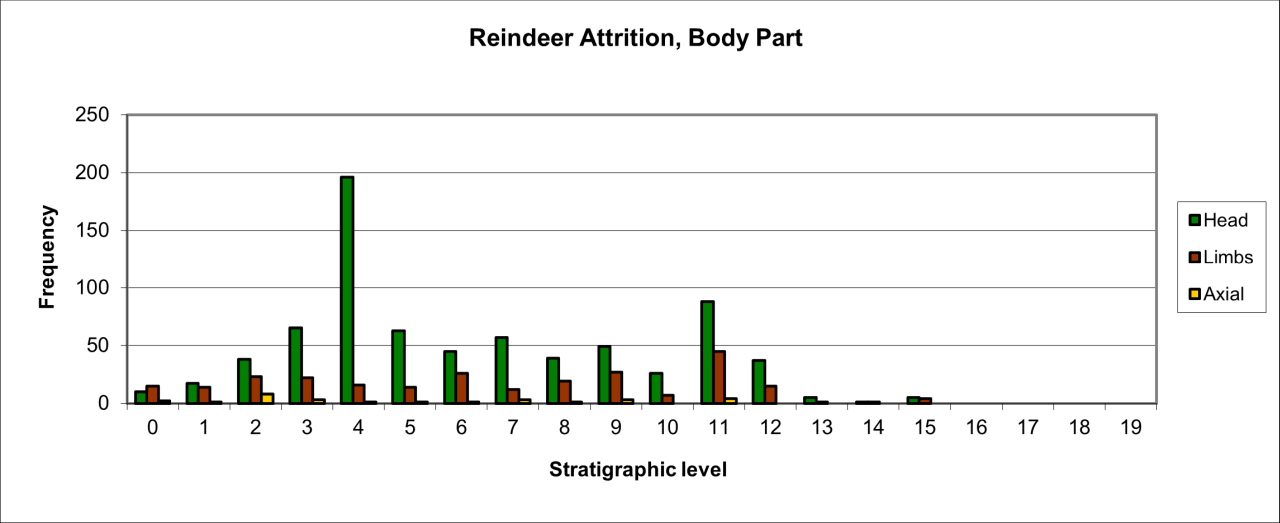
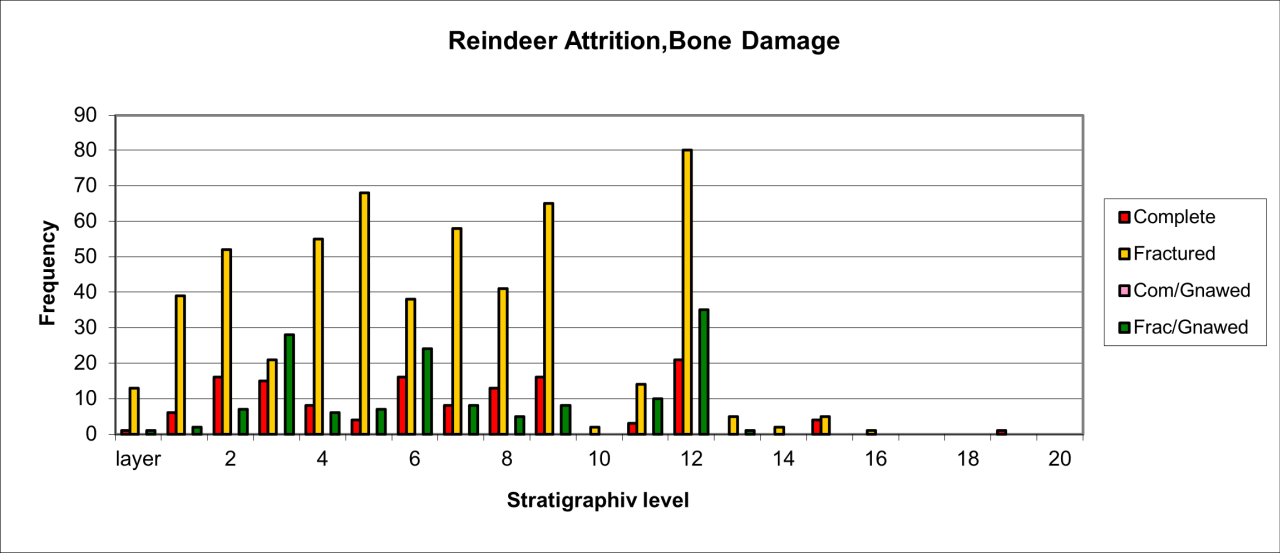

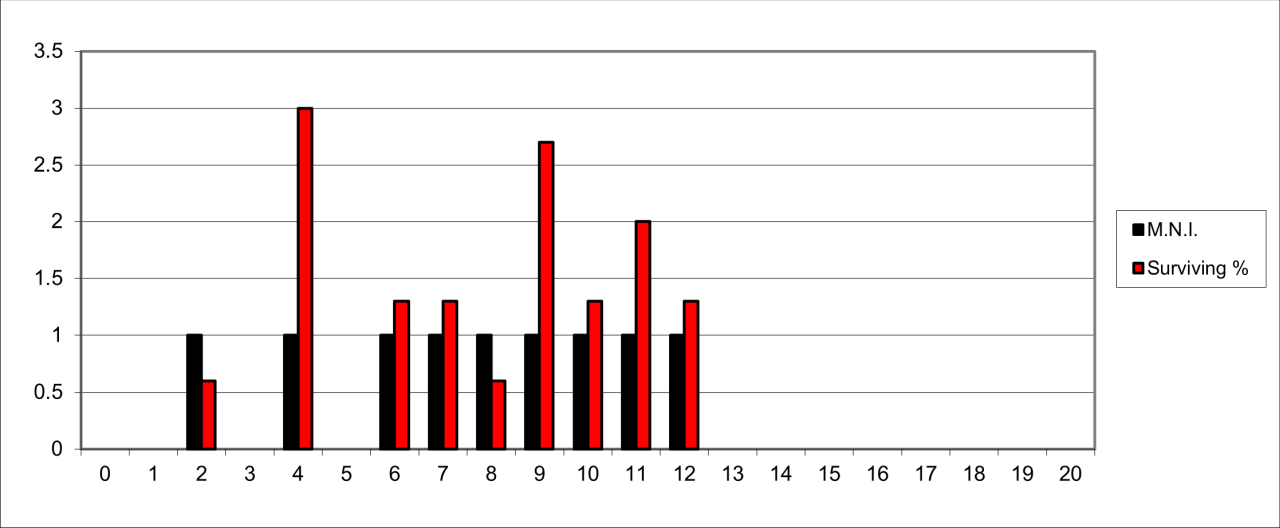
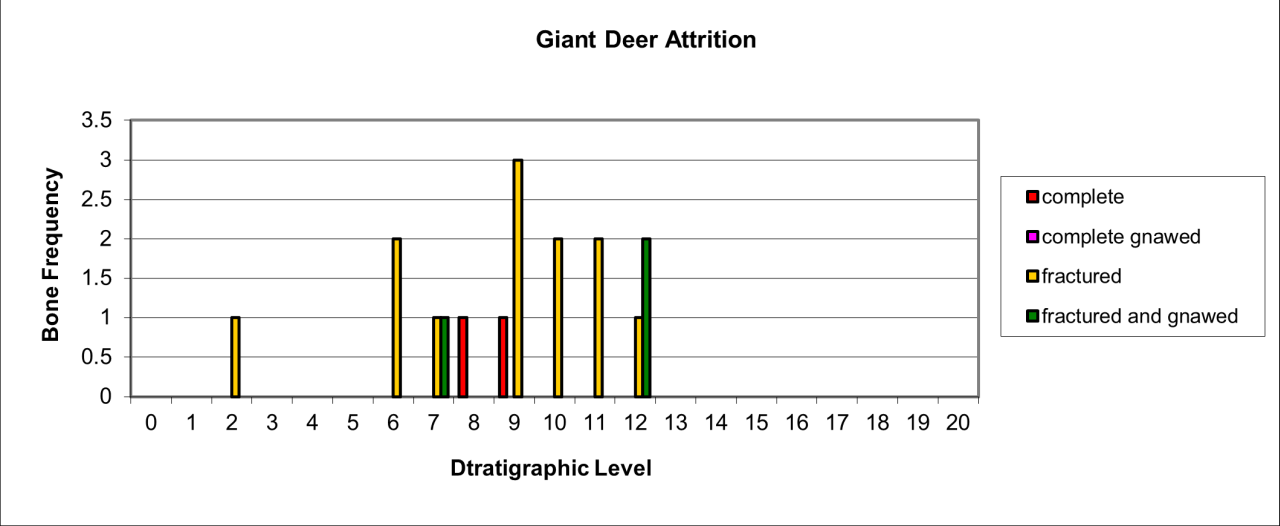
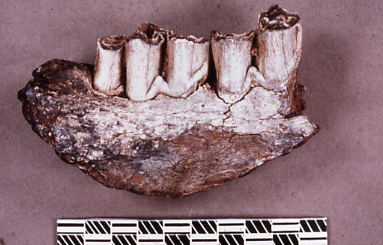
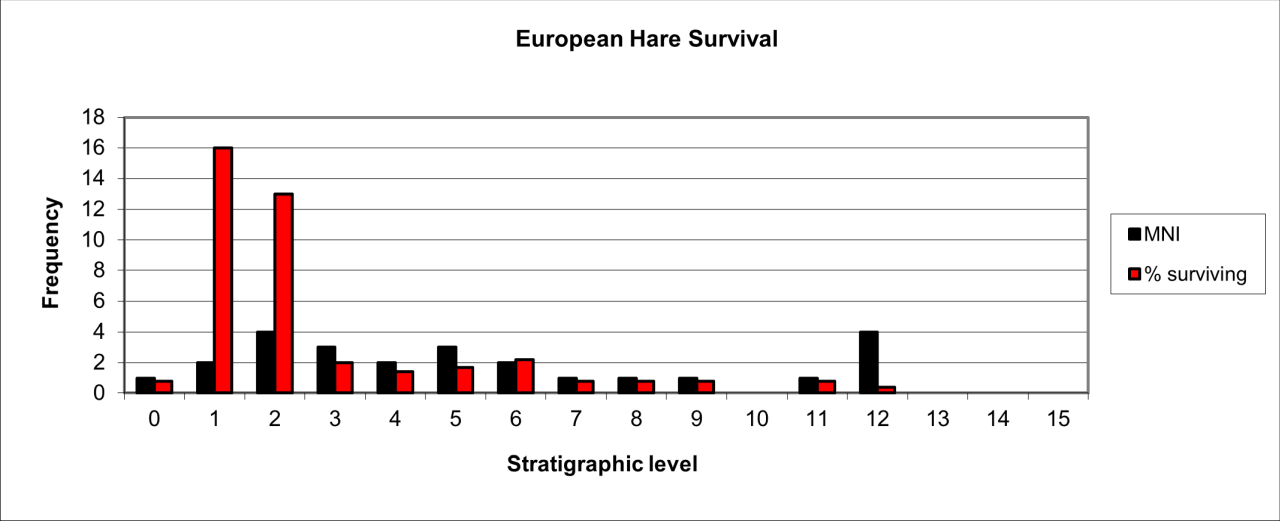
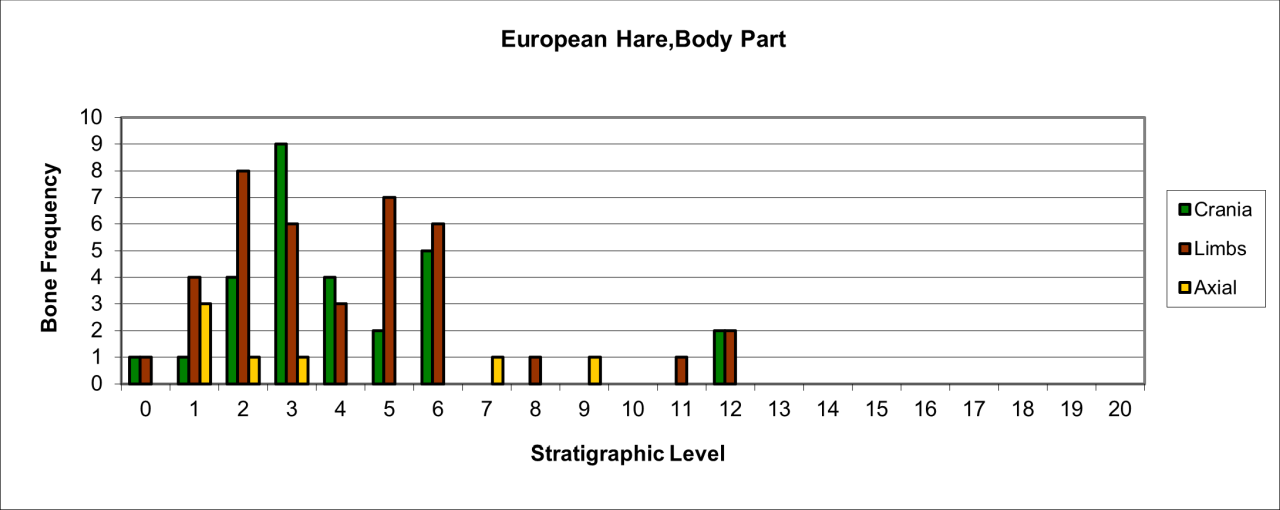
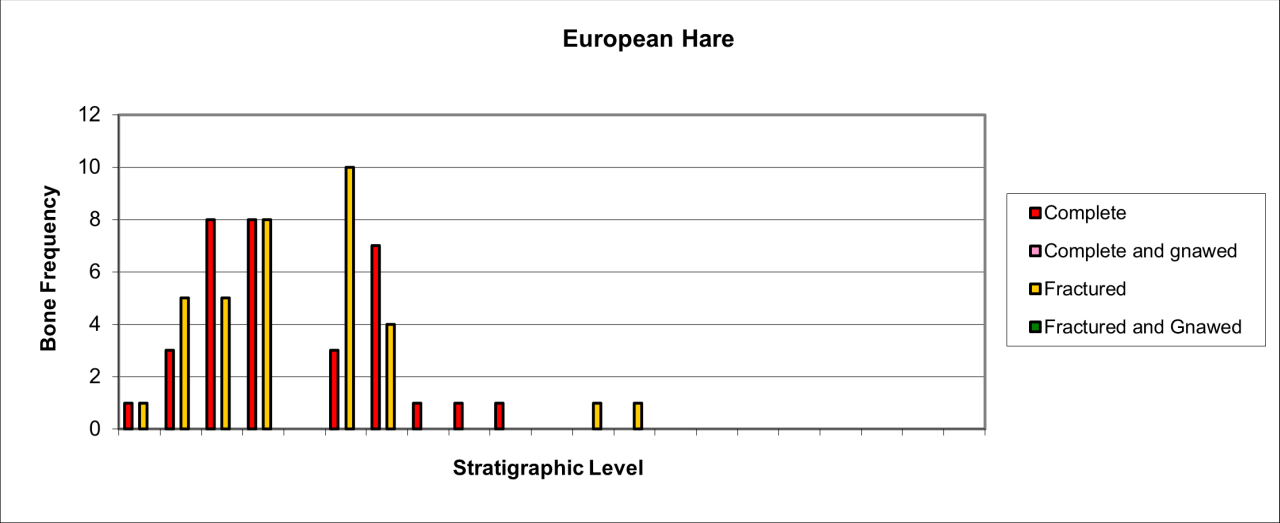
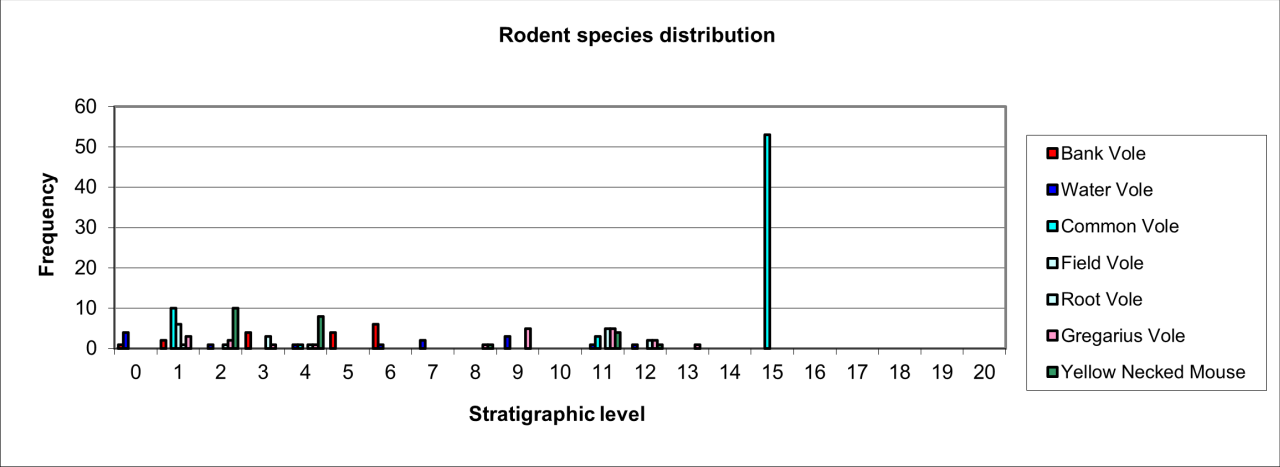
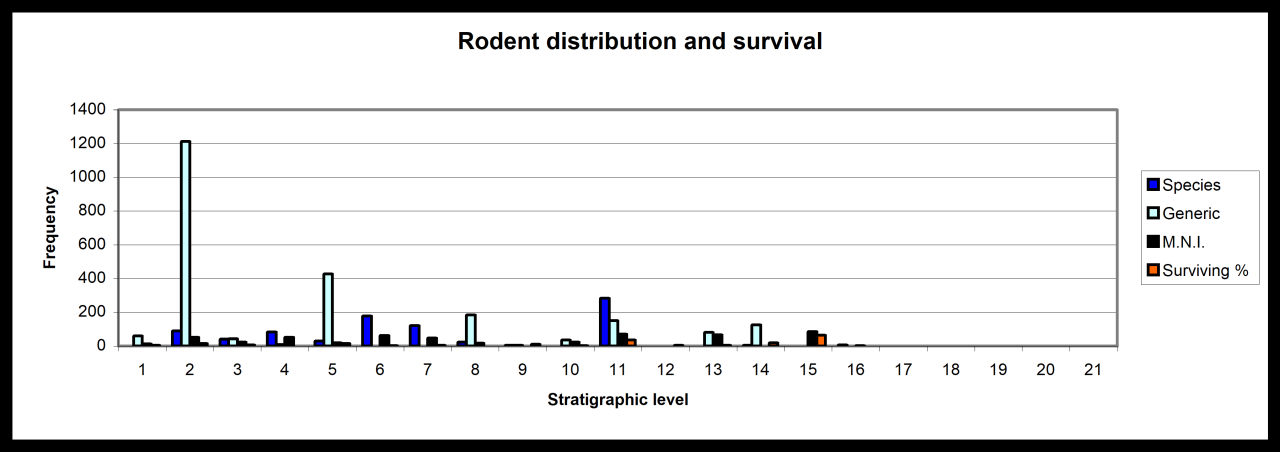
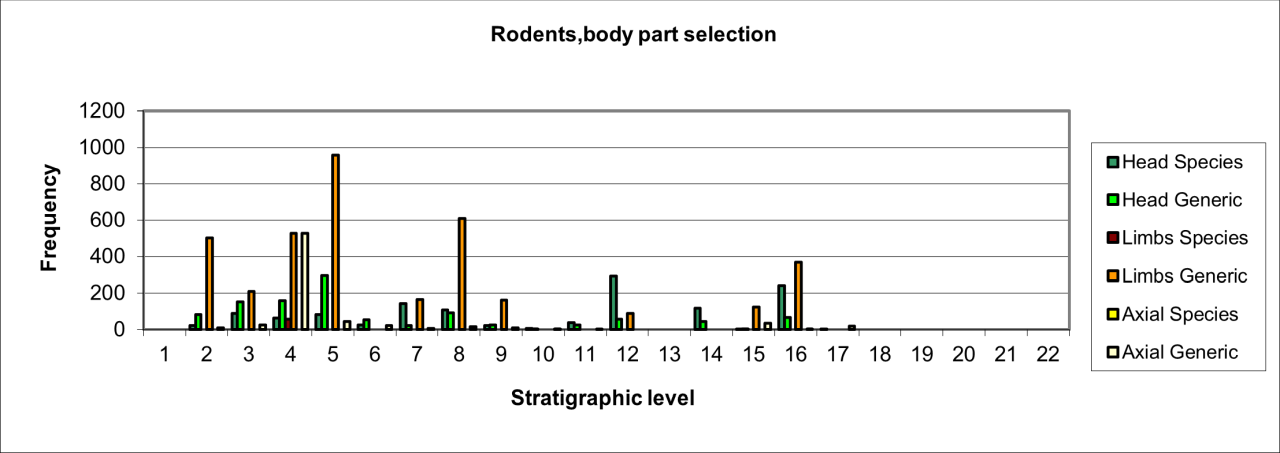
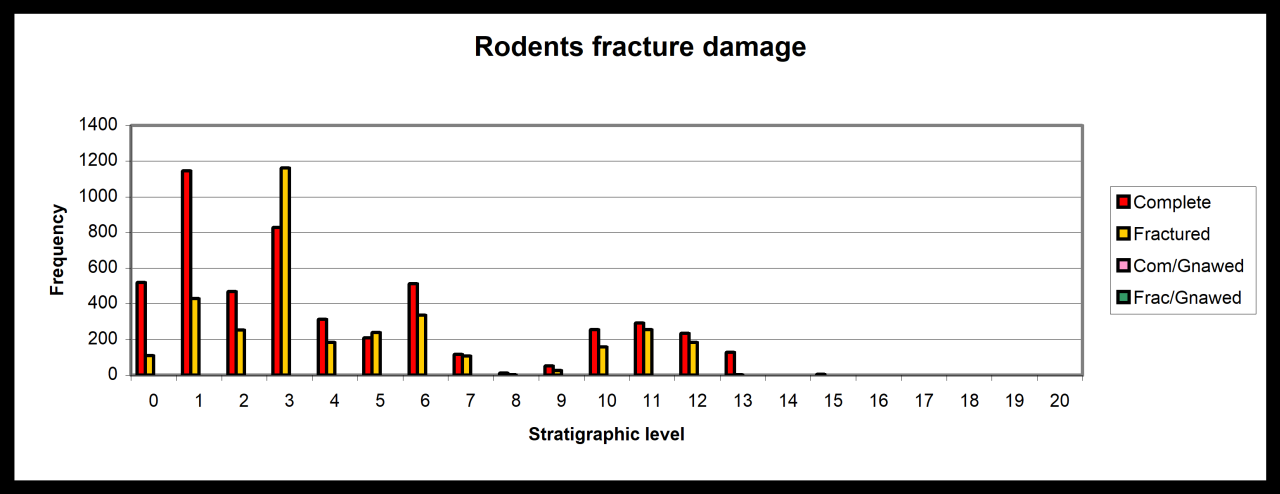
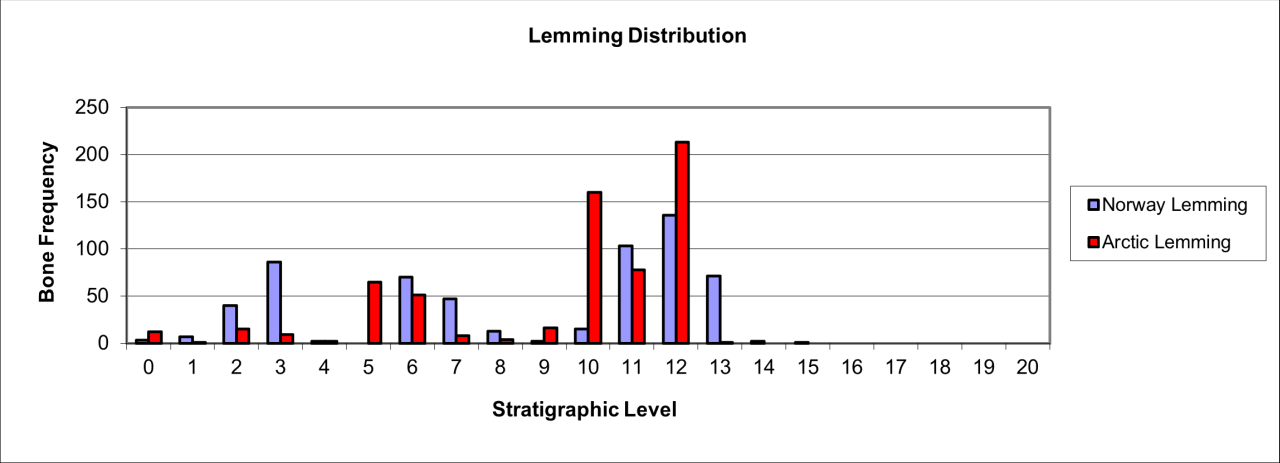
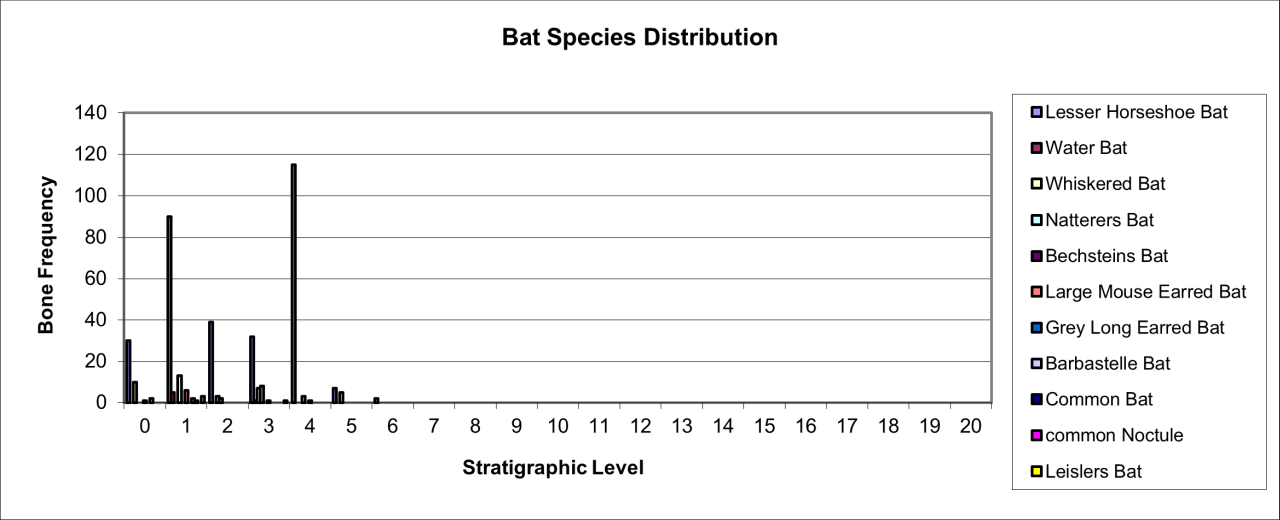
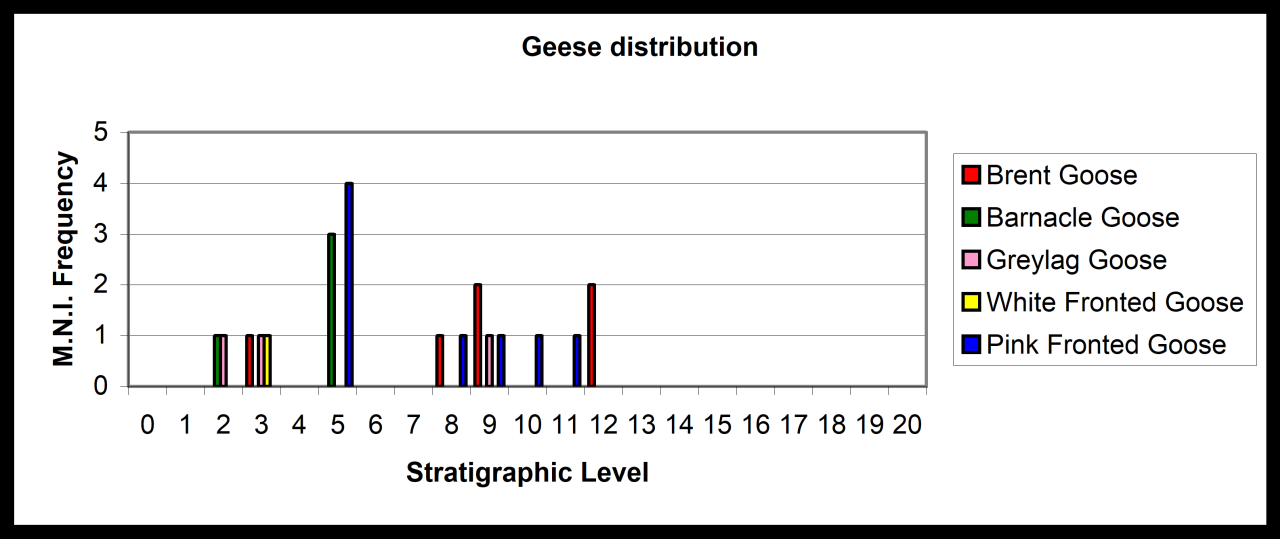
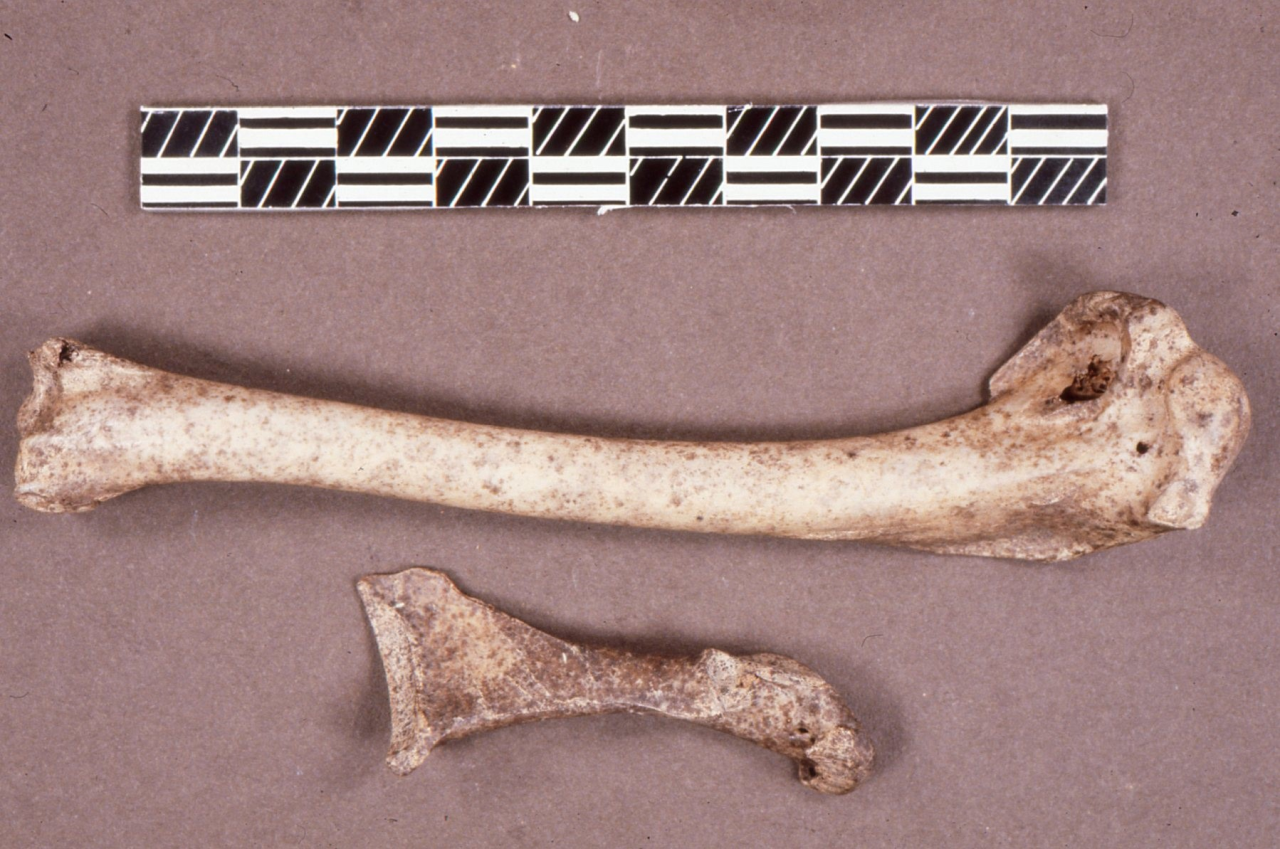



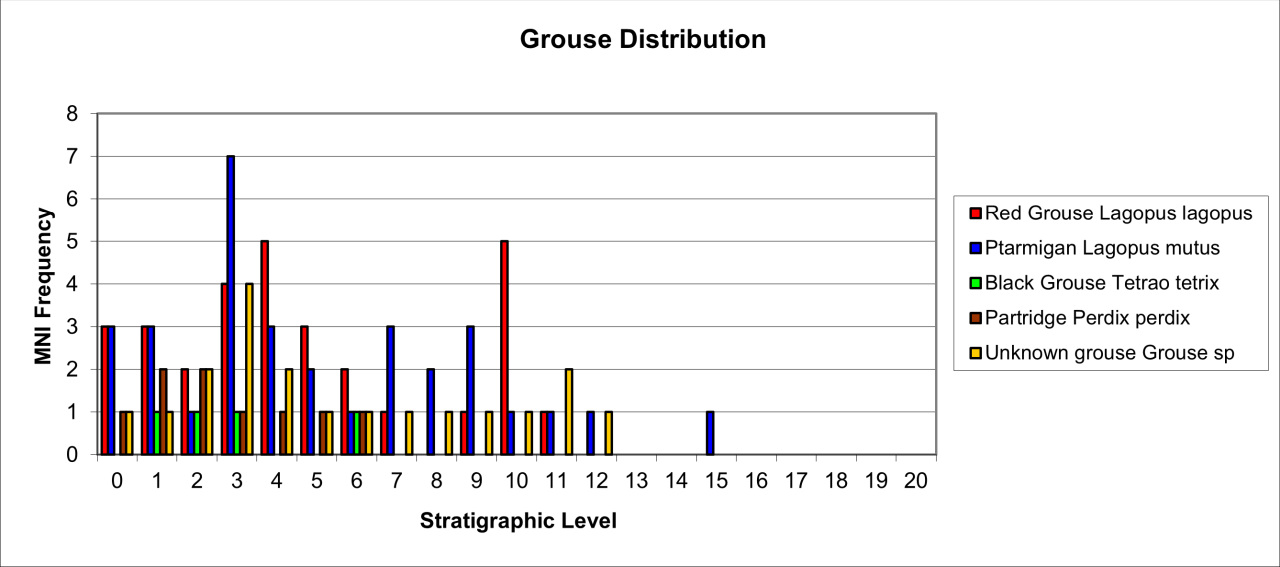
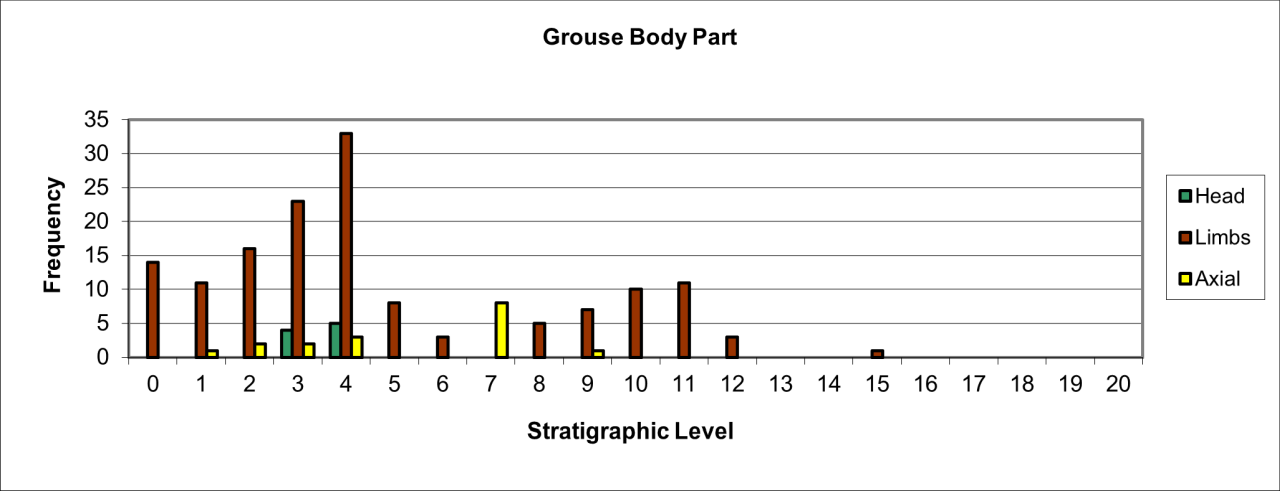
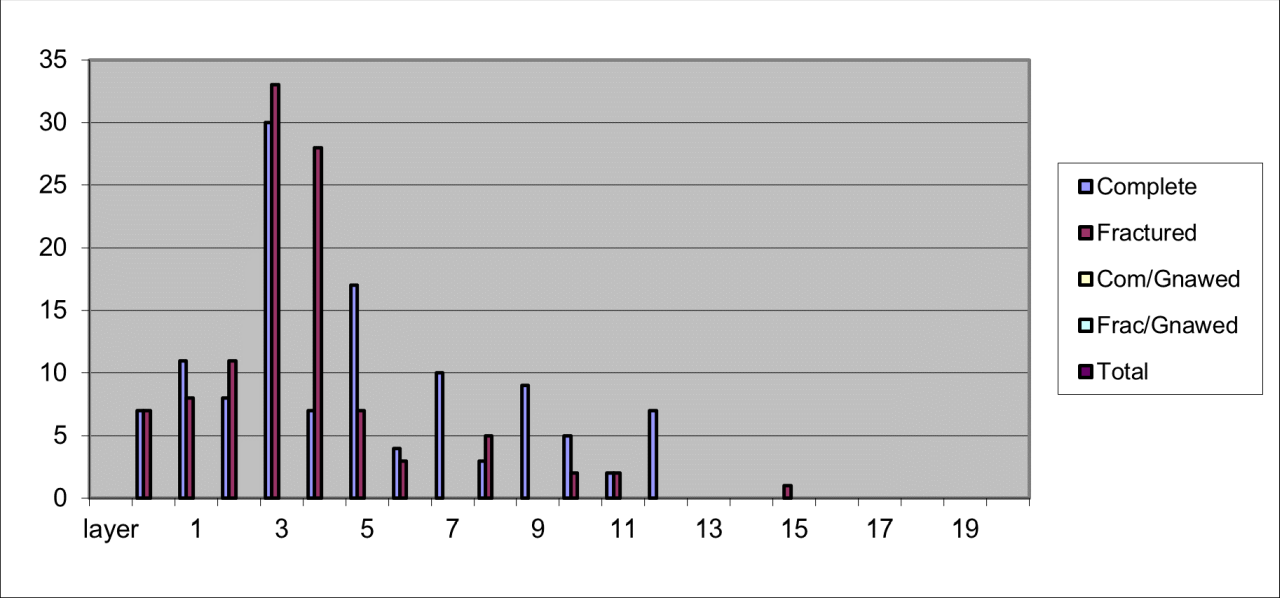
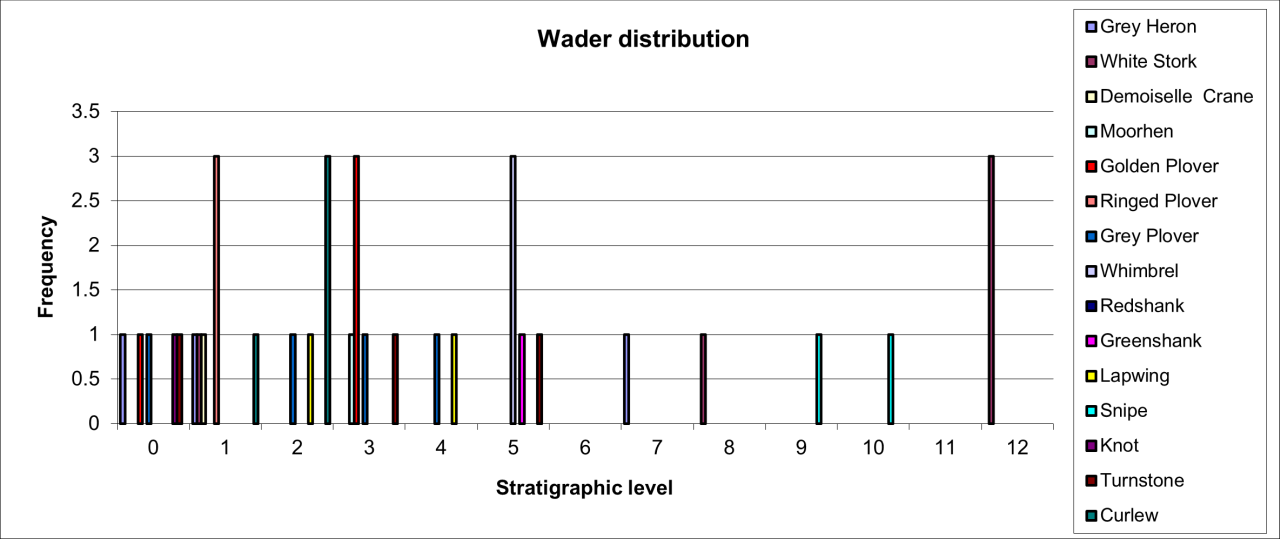
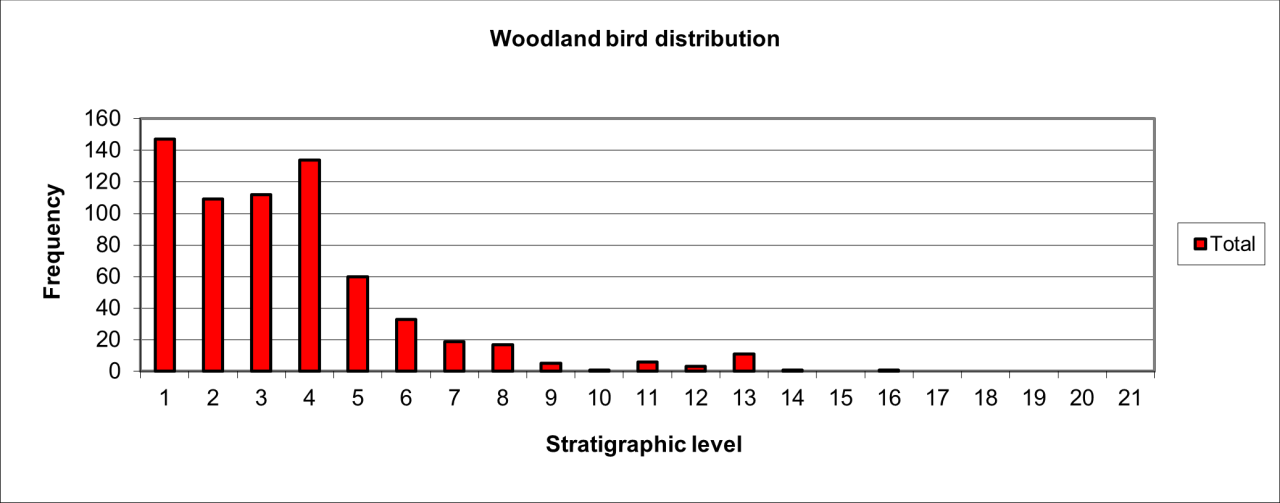
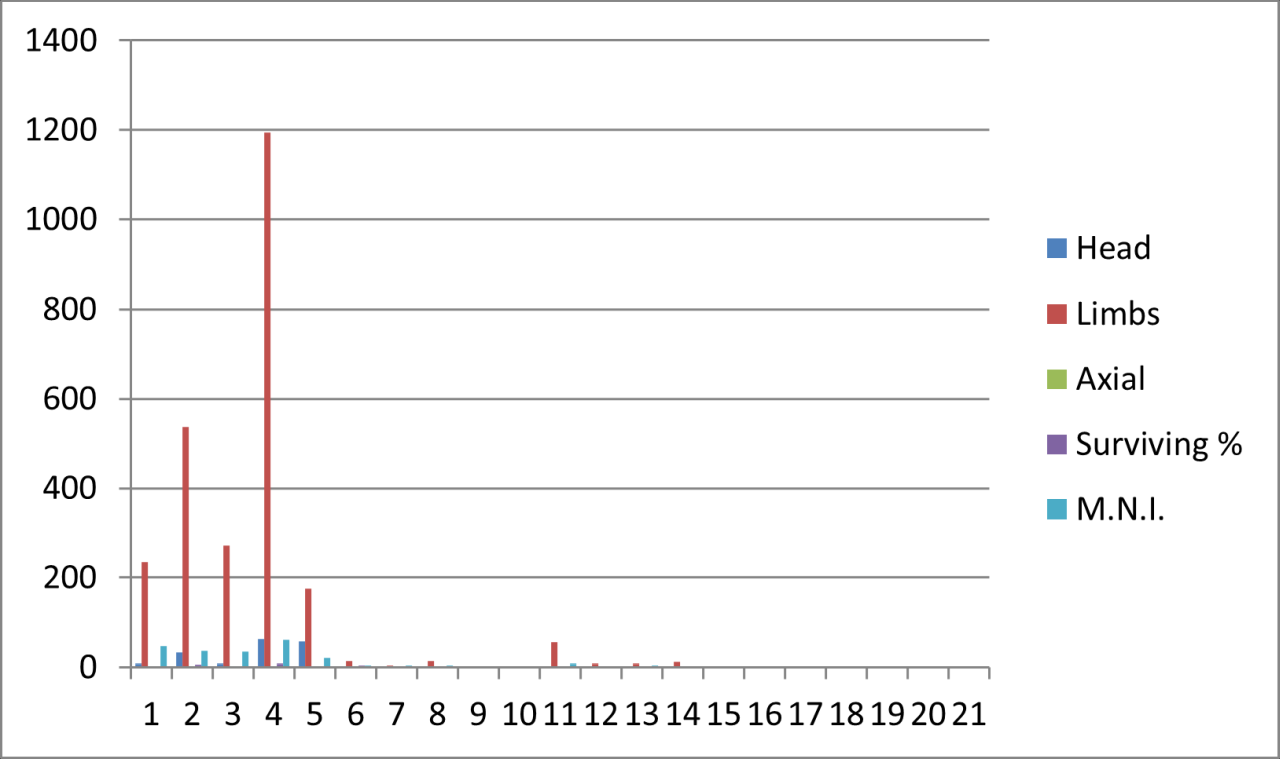
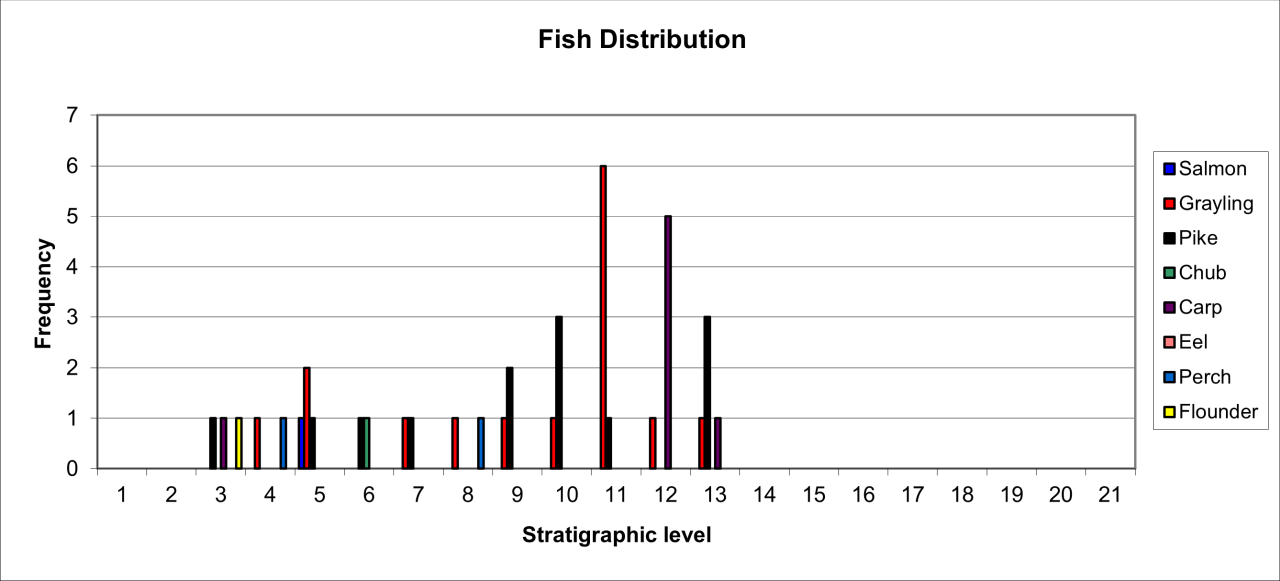
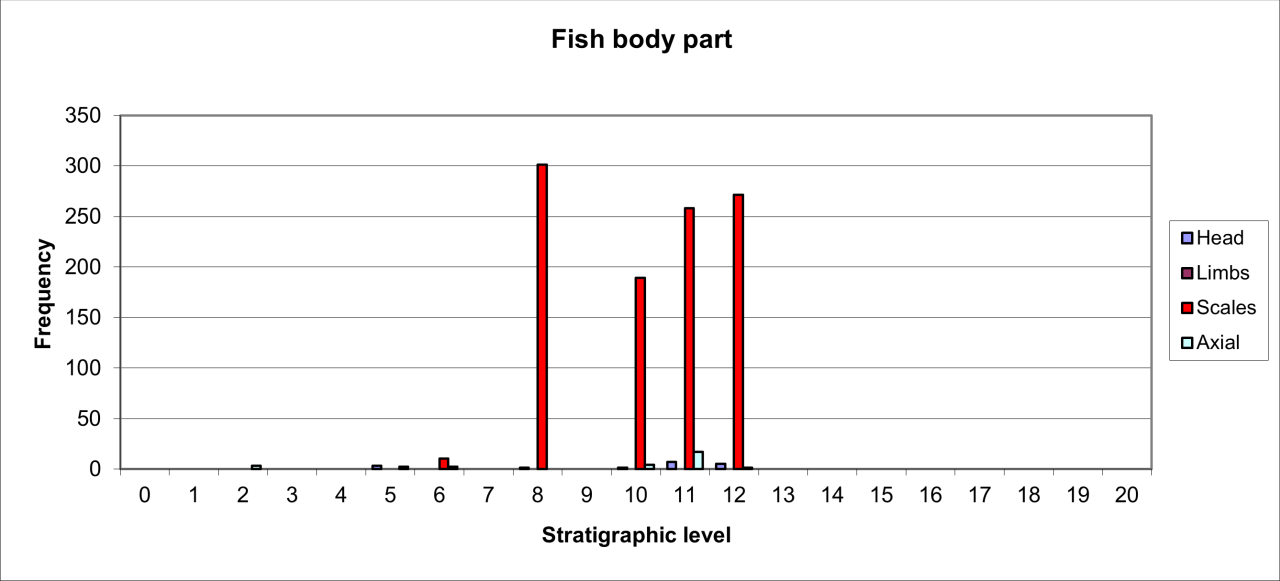
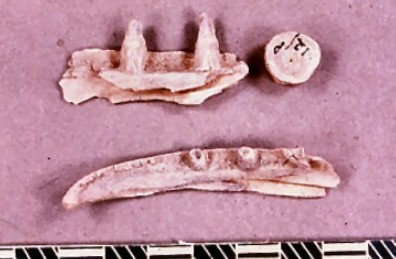
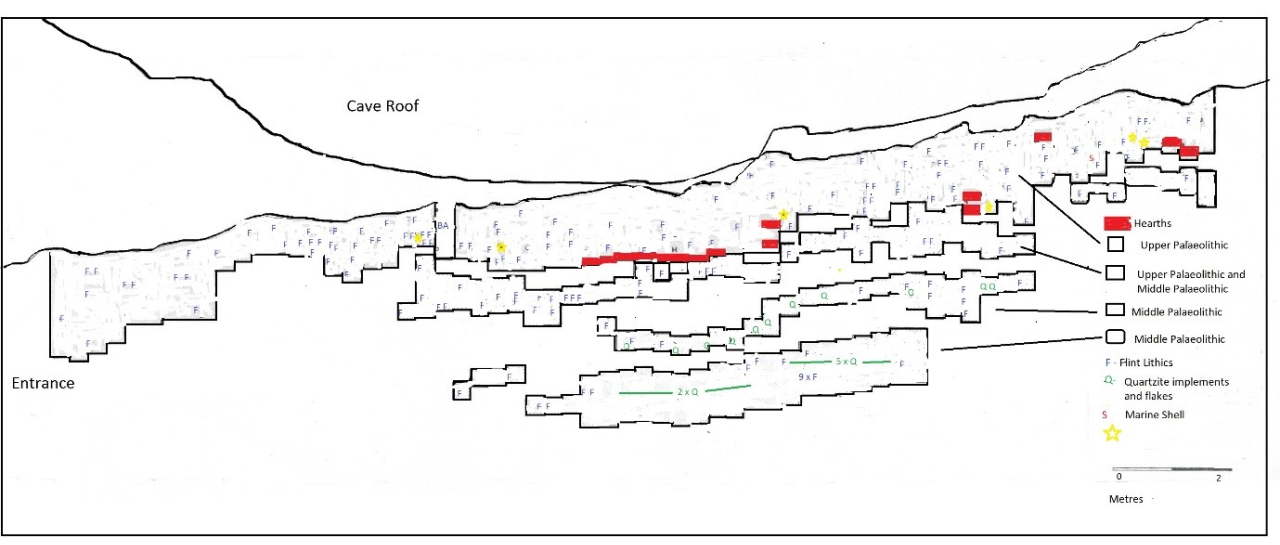{kind=link}