A North-Western Habitat: the Paleoethology and Colonisation of a European Peninsula
Rogan D.S. Jenkinson
Cite this as: Jenkinson, R.D.S. 2023 A North-Western Habitat: the Paleoethology and Colonisation of a European Peninsula (a comprehensive analysis of excavations in Pin Hole Cave, Creswell Crags), Internet Archaeology 61 (Monograph 31). https://doi.org/10.11141/ia.61.1
9. Hunting or Scavenging Behaviour
9.1 Taphonomy of the major ungulates - human or carnivore agency?
This study has attempted an exhaustive description of the Pin Hole Cave vertebrate remains, their reconstructed stratigraphic distribution, particularly in respect to the archaeological evidence, and the nature of physical damage to the preserved remains. A key issue has been to identify taphonomic process responsible for the accumulation of such a large and diverse accumulation of material and a significant aim is to try and differentiate between the predatory behaviour of humans and carnivores. In many cases, the surviving evidence is insufficiently detailed to achieve this objective.
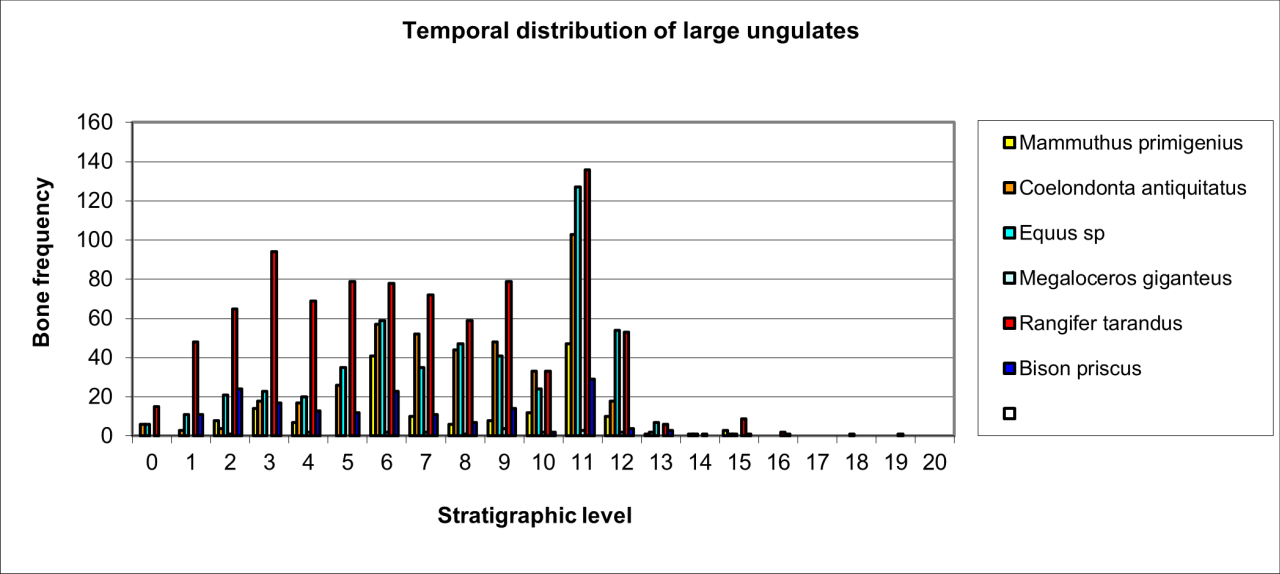
Figure 113: Summary of large ungulate temporal distribution
Figure 113 summarises the temporal distribution of the major ungulates and demonstrates considerable variation in the frequency of individual species. Reindeer, Horse and Woolly Rhinoceros are present throughout the entire sequence in considerable quantity. The remains of both Bison and Mammoth are also present in virtually all stratigraphic levels. It is quite clear that the earlier sequence within the cave contains relatively large numbers of Woolly Rhinoceros and Wild Horse that are in association with both Neanderthals and Hyaena. There is also direct evidence of human cutting of Woolly Rhinoceros (and Mammoth) bone occurring, with Hyaena-gnawed, crushed and polished bone of both species. The presence of ungnawed and/or cut bone of both species suggest that use of the cave by humans and animals is at different times. Less frequent remains of Reindeer, Bison and Mammoth also occur within the lower sequence. Numbers of the larger ungulates slowly decline until level 5 when they dramatically decrease. The upper sequence (0-4) has few large ungulates and is dominated by large numbers of Reindeer with occasional Bison. The interpretation is not comprehensive but isolates several taphonomic issues that are relevant to human occupation or use of the cave.
9.2 Human occupation. Early Middle Palaeolithic
Evidence from the lower cave sequence demonstrates that Middle Palaeolithic groups utilised the cave. They utilised, hunted or scavenged parts of juveniles of Woolly Rhinoceros and Mammoth and transported lower leg parts and mandibles into the cave rear. The lack of carnivore damage to these vertebrate bones clearly indicates that Middle Palaeolithic use of the cave was not contemporary with Hyaena denning.
European Bison remains are heavily damaged by fracturing, include axial skeletal parts and also carry little evidence of carnivore damage, which suggests that this species was an important element of human procurement. The observation that vertebrate bone with direct evidence of human use belongs to juvenile animals may indicate that there was a preference for hunting these species in a relatively carnivore-free environment and that scavenging was of little importance.
The evidence of incised bones associated with a lithic assemblage dominated by cutting edges suggests that skinning and pelt preparation were important activities within the cave.
Temporal definition within the distribution of Middle Palaeolithic lithics is not completely clear but does suggest an early Middle Palaeolithic use where tools are dominated by cutting edges fabricated from locally available 'Bunter' quartzites. This early occurrence is separated stratigraphically from later Middle Palaeolithic assemblages, which include a greater number of tools that are not only often fabricated from flint but continue to be dominated by similar tool types.
Later Middle Palaeolithic use of the cave occurs in areas where there was earlier use and suggests that the cave entrance was too wet - an observation confirmed by water-orientated bone and calcrete formations that have been discovered within this area. It is interesting to note that this is extremely narrow and the infilling floor sediments were sharply inclined toward the entrance.
The artefacts occur in stratigraphic levels which indicate a significant decline in both the remains of Hyaena and in indications of denning and gnawing damage to other vertebrate bone. The same stratigraphic levels indicate an increase in evidence of European Wolf and Reindeer. European Bison remains continue to be a significant element. This seems to indicate an increase and diversification in the procurement of resources using Reindeer and Bison and which are associated with a decline in the numbers of Hyaena, Woolly Rhinoceros and Mammoth. The role of European Wolf, a major predator of Reindeer, is enigmatic and circumstantial evidence suggests not only coeval use of the cave but also the possibility of a cooperative relationship.
The remains of Wild Horse are derived from older females, a factor that may have helped in the predation of this otherwise highly mobile and aggressive species. Many of the surviving bones are carnivore damaged and there is no direct indication of human interest.
Multiple lines of evidence that include the presence of young and cast deer antlers indicate that occupation was during spring and summer and that at this time these may have been collected as a raw material. Many are also gnawed, presumably by Wolves, and further indicate that human and Wolves are coeval.
The evidence suggests that these groups were nomadic with a seasonal (spring, summer) use of the cave and that these were widely separated in time. In both cases, there is an interdependent relationship with the movement and use of migratory species.
Within this period of cave use, Brown Bears are periodically giving birth and semi-hibernating. During winter, one very large individual may have been deliberately slaughtered.
9.3 Human occupation. Later Middle Palaeolithic
Two stratigraphically discrete lithic assemblages occur in levels 6 to 9 and the115 artefacts that can be re-plotted are stratigraphically separate from the earlier assemblage. Many of the lithics are clearly marked with location details, and include both flint and quartzite raw materials. Both assemblages extend along the inclined cave floor from the passage into the 'inner chamber'. There is considerable diversity in typological form and raw materials. The assemblage is dominated by bifaces of quartzite, scrapers and points, which are associated with broken tool fragments and fabrication waste. Considerable discussion has taken place concerning the typological variation of artefacts (Mellars 1974; Campbell 1977; Jenkinson 1984; Coulson 1990; Jacobi et al. 1998; White and Pettitt 2011), particularly in respect of well-known French and European sites. It has been suggested that the assemblage is 'mixed' or multiphase, and in any case broadly similar to the Mousterian of Acheulean Tradition. The stratigraphic location of a large invasively retouched flake, normally present in Upper Palaeolithic assemblages, has been called into question. Before reaching any conclusions it is worth re-stating that the assemblage components are well-recorded by a very competent excavator and are stratigraphically discrete in occurrence.
Within this study, artefacts from the mid-sequence, which were referred to as Early Upper Palaeolithic by Campbell (1969) are considered as Middle Palaeolithic. Despite the known effects of fluvial action in some parts of the cave. it is impractical to argue that downward displacement has taken place, which in some cases would had have to be specific and up to 1m and up slope! As a consequence, despite inclusion of pieces that are distinct (typologically and perhaps chronologically), which has implied both temporal and geographic influences from elsewhere, the assemblage should be considered as a unit and overall of Middle Palaeolithic form.
Middle Palaeolithic artefacts are associated with large quantities of palaeontological remains. The four stratigraphic levels have produced 3922 vertebrate bones belonging to a wide range of species, which in addition to large carnivores and ungulates include ground-dwelling rodents, ducks, geese, cranes and fish. This is a numerically rich and diverse animal population. Analysis of the remains from these levels suggests:
The lithic assemblage is well recorded and is unusual within the Creswell sites in that a diverse range of types are clearly associated stratigraphically. This is more clear and distinct than other poorly excavated sites at Robin Hood's and Church Hole Caves.
A relatively wide range of tool types are present, which includes scrapers and cutting edges in association with broken tools, cores and fabrication waste. Many of the tools are in pristine condition with little evidence of use. Although the collection in total is small by European standards it does, perhaps, indicate a more intense use of the sloping cave rear.
The ungulate population with all body parts, including axial parts, are now represented and the species Wild Horse and Reindeer are dominant. Most of the specimens are complete or simply fractured. The characteristic gnawing and fragmentation of bone is now rare but smaller quantities of gnawed bone are present in each level. The presence of specific species, body part selection and bone attrition state differs from the earlier carnivore accumulation. This strongly suggests that human groups are the main accumulator. A large number of Reindeer cranial parts include cast antlers and may indicate deliberate collection of these for use as a raw material.
This is a time of change in the composition of the carnivore populations. There is a very dramatic rise in the frequency of Wolf, the presence of which increases through time, and in association with similar increases in human presence. The species is obviously responsible for much of the gnawed bone, including cast Reindeer antler and the gnawing of axial body parts, a situation that is not common in earlier carnivore accumulations This suggests that these species were hunted by human groups in a context where their control and access at kill sites to carcasses ensured selection of most body parts with little competition from large carnivores. The style of occurrence and preservation of Wolf within the cave suggest that they were a live population and not hunted as a resource for human groups (i.e. skins). The seemingly close association of this species and the nature of the vertebrate bone accumulation suggest at least a mutual relationship with human groups that may have been cooperative.
The presence of diverse Duck, Wader, Crane and freshwater fish remains suggest a local environment with abundant shallow freshwater resources. A small occurrence of woodland bird species suggests that areas of scrub or woodland existed in and around the Creswell Gorge. All species indicate a spring or summer presence where there is ready access to open water and vegetation. The species presence within the cave obviously results from them being prey. There are isolated pieces of evidence indicating predation by small carnivores (Red Fox bite marks) and where multiple fish scales have been found stratigraphically associated with European Brown Bear.
9.4 Regional Middle Palaeolithic localities
Evidence for Middle Palaeolithic activity is both rare and scarce within the eastern area of the region. Significant discoveries have been made at Lynford Quarry which was discovered in 2002 (Boismier et al. 2003; Cooper 2004). At this locality, a series of fluvial sediments revealed a lithic assemblage of Mousterian type which is dated to MIS 3 (Boismier et al. 2016). The assemblage is predominantly composed of complete fabricated artefacts and there is little evidence of tool manufacture on site. It is associated with 1345 vertebrate bones, 91% of which have been identified as Mammoth (Mammuthus primigenius). Studies of the taphonomy by Schreve (2006) have reported that 67 vertebrate bones are carnivore damaged, most probably by Hyaena and these were associated with the bone of 8 Reindeer, 6 Woolly Rhinoceros (Coleondonta antiquitatis) and Horse (Equus sp). Much of the vertebrate bone is damaged or fractured, which is thought to be the result of trampling by ungulates. There is no evidence of surface cut-marks on the vertebrate bone and no direct association with the lithics found nearby. Schreve has noted that the vertebrate population, particularly Mammoth, is represented by cranial and axial parts and a significant absence of limb bones, suggesting that Neanderthal groups may have removed the available and more portable limbs as a meat resource (Schreve 2006). There is no clear evidence explaining the presence of this species in these circumstances. Tthe locality may have been a butchering site where Mammoth may have been driven into marshy ground, disabled and killed but while this is a possibility, it is not known to be a common strategy used by carnivores. It is equally likely that this indicated exploitation of natural fatalities, which are frequently located in aquatic environments. The frequency of artefacts may simply indicate repeated use at a well-known 'water hole'. The locality is clearly of some significance and the style of vertebrate bone preservation contrasts with the comparative vertebrate assemblages known from the Steetley Cave small carnivore den, the fluvial Hyaena scavenging activity known from Fulbeck and the taphonomy of vertebrates from Middle Palaeolithic vertebrates known from Pin Hole Cave.
There are several observations that are relevant for an understanding of the Pin Hole evidence. Firstly, there is direct evidence from the cave that Neanderthal groups were interested in the same species (Mammoth, Woolly Rhinoceros and Reindeer) as resources. In Pin Hole Cave, the bone remains are of juvenile individuals and have surface cut-marks. There are two issues concerning the species. Firstly they are migratory, probably over long distances, which suggests that the site is not only seasonal but may have an opportunist aspect. Secondly, little is known of Neanderthal ability to hunt and kill large, mobile and potentially aggressive species. Such actions seem improbable for groups of individuals and may have only been feasible for juvenile, isolated, infirm or trapped individuals. At both Linford Quarry and Pin Hole Cave, Mammoth remains are often of axial and cranial parts with a relative absence of limbs, which suggest that limbs were scavenged as a source of meat and that other body parts were difficult to access. While absence of evidence is not evidence, the relative lack of limb bones of Mammoth suggest that the legs could have been segmented for removal. Such a process would have involved the difficult task of cutting or fracturing of the medial leg epiphysis and the much easier separation from shoulder blades where segmentation involves little cutting of flesh.
The unusual evidence of incised tibia of Mammoth and Woolly Rhinoceros from Pin Hole Cave offers direct evidence of Neanderthal interest in the limbs of both species and which have been transported away from a death site. In both cases the remains are derived from juvenile individuals. The same locality, in levels with Middle Palaeolithic lithics, has also produced an antler fragment of Reindeer (Rangifer tarandus) that has a partially burnt tine, which may suggest that interest in this species as a resource is not necessarily related to food procurement. Analysis of the presence and condition of skeletal parts at Pin Hole suggest that Neanderthal groups utilised body parts of Mammoth, Woolly Rhinoceros, Reindeer and European Bison as resources. The evidence from Lynford Quarry offers correlative evidence for the scavenging of the larger of these species within local environments in that area adjacent to fluvial habitats.
9.5 Later Upper Palaeolithic
The dramatic change in character within the upper stratigraphic levels (0 to 4) marks the appearance of the material remains of the Upper Palaeolithic. These stratigraphic levels contain a significant increase in both archaeological and palaeontological remains. The levels have produced 138 lithics, 10 bone artefacts, 2 Jurassic fossils, 4 non-local mineral fragments and these are associated with 14,556 vertebrate bones that derive from, at least, 1054 individuals and which include multiple examples of cut and incised antler and ivory. The available data suggest a range of observations, which also demonstrate a significant change in the archaeological record within the cave. These are:
Human use of the cave extends over the whole length of the cave including the small inner passage previously unused.
There are at least two areas of charcoal concentration within the rear of the cave, which were interpreted by Armstrong as the remains of hearths. The surviving charcoal includes fragments of wood.
The evidence from tool assemblages indicates a complete change in production technology and the extensively recorded switch to flint blades and their adaptation into more complex tool types. For the first time, the range of tools indicate a diversification in functions performed, and while the assemblage retains frequent cutting edges, in the form of sharp blades, these are accompanied by points, awls and piercing tools of bone and flint and a multiple range of scrapers.
Bone artefacts make their first appearance in the form of rods of ivory, bone and antler and some of these are incised in at least three different 'designs'. Their function is unclear but the nature of their design suggests that they may be parts of projectiles.
An ambiguous curiosity associated with these groups is demonstrated by the presence of a Jurrasic Echinoid and Belemnite, possibly from deposits to the east in Lincolnshire, a fragment of shell from a marine species and an amber nodule. It is not clear if these were of decorative value or curiosities. It is also possible that older pieces of tusk or ivory found their way into the later levels in this way.
Cast antlers of Reindeer are prolific and outnumber the minimum number of individuals suggested by that of the species' post-cranial bones. This clearly demonstrates that cast antler of this species was collected and cached as a raw material within the cave.
Both Reindeer and European Bison initially increase in frequency within these levels and are represented by all body parts, including axial parts, many of which are either complete or have been simply fractured. Gnawed bones are present within the earlier levels but their occurrence diminishes. The only large predator known from these levels is Wolf, which is likely to be responsible for gnawing damage.
A considerable rise in the frequency of Wolf characterises these levels and they exhibit a fairly high survival rate represented by all body parts in a complete or simply fractured form. Despite there being no systematic study, it is clear that young and juvenile individuals are present. There is no clear indication if their presence was the result of predation by human groups, as a food or pelt resource, or if they may have been in a cooperative relationship with human groups. Their sudden decline in numbers within the uppermost levels is enigmatic.
The evidence for the Upper Palaeolithic is associated stratigraphically with many other 'prey' species but there is no direct evidence of human predation and in fact the style of preservation and attrition suggest an accumulation by Wolf, smaller carnivores and raptor bird species.
It is clear that very little of the five species have been preserved as skeletal remains within the cave. The available evidence shows that less than 10%, often much less, of the carcass is available from the original vertebrate population. Unidentified bone and small, difficult to identify fragments, while numerous, would not significantly alter the overall quantities indicated. In such circumstances the interpretation of taphonomic process requires some caution. It is, however, more clear that the very high quantities of missing skeletal parts suggest that complete carcass remains were never transported into the cave and that the individual animals have been dismembered, butchered or scavenged elsewhere and outside of the cave environment. The assemblage therefore reflects a highly selected sample and it is very significant that very few axial parts are preserved within the cave. These skeletal parts include ribs, vertebrae (including centrums) scapula and pelvis. These are not fragile bones. While the parts themselves are flesh covered, more importantly they encase the major organs and abdominal area. Variation in the preservation of remains is of the same order of magnitude for each species, where an average of 90% of the original live skeleton is absent or destroyed elsewhere.
Available edible food resources reflected by the carcass weights of surviving individuals vary considerably. There are relative peaks in quantity for Mammoth and Woolly Rhinoceros within stratigraphic levels 11 and 12. These are associated with both cut bones and carnivore-damaged bone where there is evidence of stratigraphic separation of each groups within each level. There are also considerable peaks in the occurrence of Reindeer as an edible food resource within the upper stratigraphic levels and which is associated with both Middle and Upper Palaeolithic evidence.
The representation of body parts within the overall context of selection and destruction for all species is highly variable for each stratigraphic level and suggests a number of causal factors. Mammoth and Woolly Rhinoceros remains surviving within the cave are derived from a low number of individuals but present a considerable food resource. Both species are represented by relatively high number of cranial and limb pieces, which are in many cases apparently derived from single individuals. In addition to the physical difficulties of killing individuals of either species, the selected fragments would have been some of the heaviest and difficult to transport pieces. The relative lack of axial parts, which are in any case made up of large bone fragments, is also of interest and is an issue discussed below.
In stark contrast, there is a significant presence of Reindeer (as edible food) within the upper levels and associated with Upper Palaeolithic presence within the cave, and selection is much more diverse in a context where Wolf is present but Hyaena is absent. The temporal variation in body part survival, which is accompanied by a decline in Hyaena and its replacement by European Wolf, and the initial presence of Neanderthals replaced by Upper Palaeolithic groups in the later sequence, suggest considerable change in the nature of food and resource procurement.
9.6 Large ungulate population - the occurrence and condition
The clear identification of relationships between human groups and the vertebrate population is one of the main objectives of this study. Analysis of the archaeological and palaeontological evidence in total and the context of Armstrong's excavation have demonstrated a number of key issues relevant to understanding the species' relationships and context of occupation and dispersal. Despite the relative care exercised within the original excavation and a definition to 0.3m of sedimentary infill, there are limits. A 0.3m level of sediment must represent a considerable period of time during the Quaternary and this no doubt contains a multiple range of episodic events that are difficult to individually detect and separate stratigraphically.
The much-cited occurrence of Hyaena-crushed and gnawed vertebrate bone, while an important taphonomic element, is confined to a narrow range of large species. When considering the bone damage and attrition in the context of the total assemblage, carnivore crushing and gnawing can be seen to be an obvious element within the assemblage but is a secondary one. It is unfortunate that generations of researchers have been attracted to the impressive presence of Hyaena and its obvious behaviour toward its prey to the extent that the wider context of the palaeontological evidence has not been noticed.
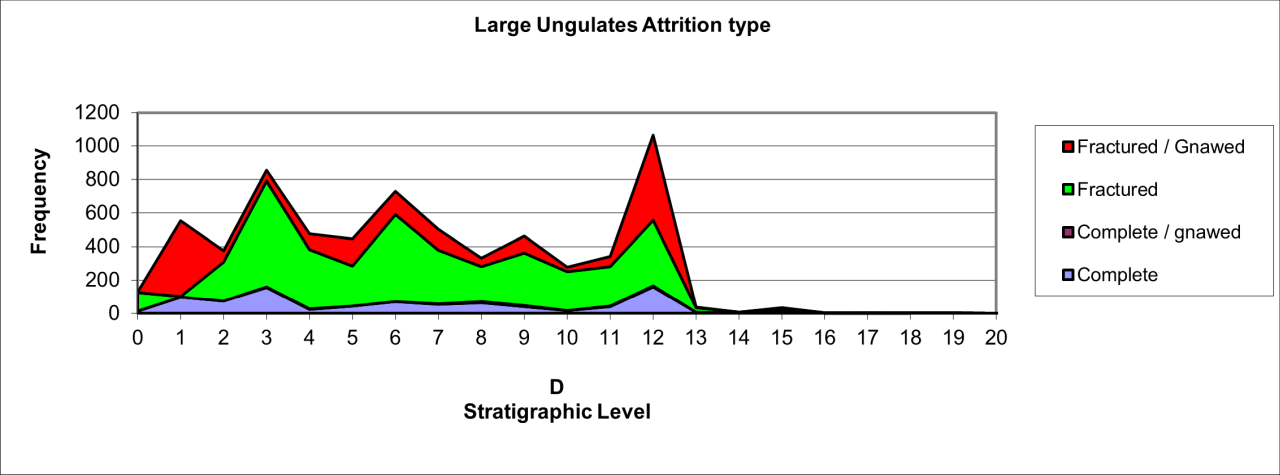
Figure 114: Temporal variation in vertebrate bone attrition
Although a massive amount of selection and reduction of individual species has taken place, the overall sequence indicates that many of the vertebrate bones that survive do so in a fairly complete condition. Much of the remaining bone is fractured and although the nature of fracturing has not been systematically recorded in this study, it is clear that there are several causal factors resulting in bone breakage. The frequency of carnivore-gnawed bone, while numerically large, is a fairly small component of the overall faunal accumulation.
The temporal occurrence of complete and fractured bone, shown in Figure 114 shows a remarkable similarity to the occurrence of human occupation indicated by the presence of lithics within the cave. The occurrence of a combination of artefacts, increased amounts of bone, cut bone and fractured bone is relatively distinct in levels 10-12, levels 5-8 during the Middle Palaeolithic and levels 1-4 for the Upper Palaeolithic. In each case, the presence of gnawed vertebrate bone is low in relation to the total remains. The long-term occurrence and specific amplifications closely match the same observation noted for human groups, Reindeer, European Bison, and European Wolf, and are different from similar observations for other large predators and ungulates. The circumstances are not individually clear and are confused by coeval residence and use but the consistent association of artefacts with increased quanties of vertebrate bone, complete and fractured bone may be diagnostic of human activity being the main taphonomic process within these levels.
9.7 Large ungulate body part frequency
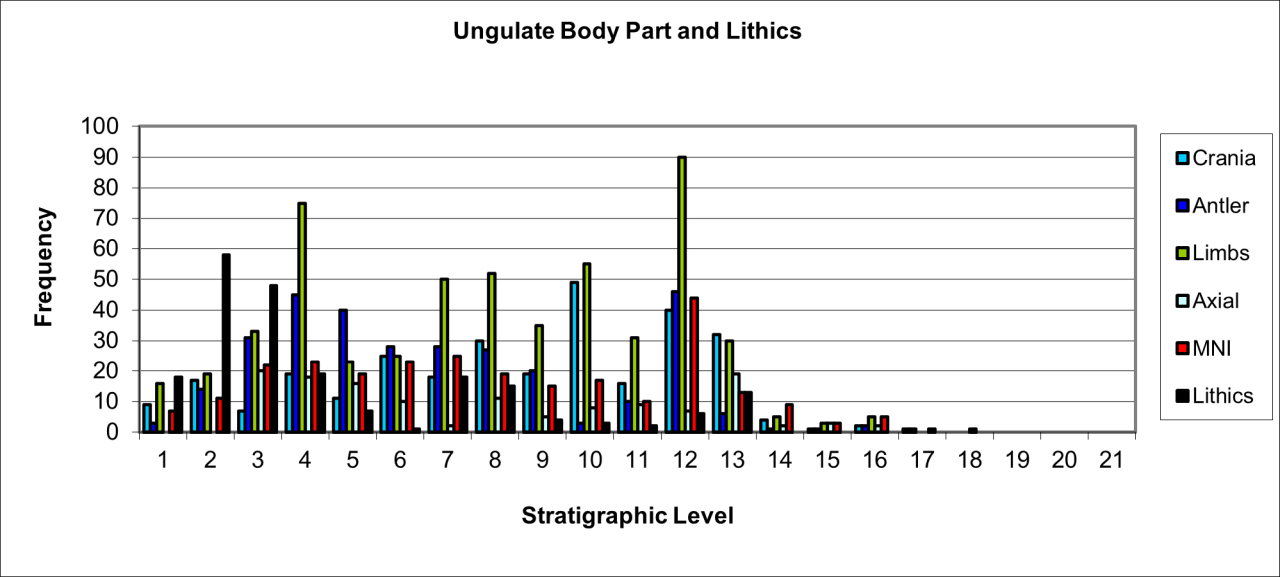
The temporal distribution of body parts for the large ungulate species, Woolly Rhinoceros, Mammoth, Wild Horse, European Bison, Reindeer and Giant Deer are shown in Figure 115. The figure attempts to show a relationship between their variation and their association with archaeological lithic evidence. This evidence has already been described and occurs separately in levels 11 and 12 for the Early Middle Palaeolithic, within levels 6-9 for a Later Middle Palaeolithic and in levels 1-5 for the Upper Palaeolithic. Hyaena denning and use of the site occurs in levels 6-12 and wolves are present in levels 2-11.
Figure 115: Large ungulate body part frequency
The frequency of ungulate body parts is dominated by cranial bones, which are shown separately as cranial bone, teeth and Reindeer antler. Post-cranial bones occur in large numbers as limb fragments associated with some axial parts. There is a distinct stratigraphic association and temporal variation of these body parts with the occurrence of lithic evidence, which suggests that human groups were a significant causal factor of ungulate bone accumulation. This is particularly distinct within the upper stratigraphic levels where large numbers of cast Reindeer antler are associated with a large number of lithics. Many of these levels are also characterised by much lower frequencies of carnivore-gnawed bone and a significant absence of Hyaena and surface salivated bone within the upper levels. Salival surface polish and scalloped bone fragments are largely confined to the early levels.
The nature and diversity of body parts clearly indicate an element of control at kill sites, outside of the cave environment, and where the lack of inter-species competition has allowed a selective removal of parts, which is motivated by preference and not opportunity. This implies a hunting strategy rather than scavenging activity. The analysis suggests that within the overall accumulation and blurred stratigraphic definition, which inhibits separation of episodic events related to both human and carnivore activity, that it is possible to separate, to some extent, these taphonomic agencies. Despite the clear lack of definition of human activity, it is apparent that the lithic assemblages that have some element of association with vertebrate fauna is of a specific type, and includes many fresh, new and unused tools. It seems that the assemblage does not include the tool types needed to process cranial and limb parts of large ungulates within the cave. This seems to indicate that much butchering and preparation of carcasses into a food resource must have occurred elsewhere.
The food resources available to both humans and vertebrate populations can be reconstructed from the skeletal remains of ungulate bones within the cave. These are set out in the digital archive.
9.8 Hunting and scavenging behaviour
The quest for food and water resources must have provided one of the main motivating forces causing dispersion into this region by both human and animal populations. Actualist studies provide an enormous quantity of information that illustrate the nature and parameters that may have operated to control behaviour by both population groups during the Quaternary.
An understanding of behaviour and inter-species relationships is fraught with difficulty due to the nature of the evidence and suggestions of taphonomic processes based upon fragmentary remains. Within this context, the separation of hunting and scavenging behaviour by of both groups is not generally clear-cut. Specific indications such as cut-marked bone and gnawed bone, which offer direct evidence of behaviour, offer important information but is limited by its occurrence and the difficulty of separating the activity within its stratigraphic context.
A general but useful approach is to consider the survival and taphonomy of large species such as Mammoth, Woolly Rhinoceros, Wild Horse, Reindeer and Bison. These species are too large to enter the cave as part of their natural behaviour and their occurrence within the sediments is, therefore, the direct result of hunting or scavenging activity. This activity has resulted in the differential selection and destruction of body parts, the scale and intensity of this process illustrated in the digital archive.
The Pin Hole Cave locality was a busy place during the Quaternary and the extensive human and vertebrate habitation and use has left a very large collection of fossilised bone. Differentiating and understanding the taphonomic process responsible for the accumulation is made difficult by the lack of detailed definition within the fossil record and the multiplicity of factors affecting individual fossils.
In order to calibrate the fossil evidence, and particularly to consider the varying size and behaviours of individual species, the available biomass (in terms of a food resource) has been reconstructed. The approach uses the construction of minimum number of individuals for any given species to quantify the total kilograms present in the source live population. The reconstruction (Figure 116) utilises the five concepts defined below:
Species MNI - the figure expresses the minimum number of individual animals, which are estimated on the basis of surviving fragments. These are shown for each species and stratigraphic level.
Survival % MNI - this is an estimate of the surviving skeletal parts within the sediments compared to those that would be expected to occur within an undamaged live population.
Missing % - the percentage expresses the number of body parts that would be expected in a living undamaged population living and dying outside of the cave minus the surviving parts known from the cave sediment.
Total kilos of edible food - the total amount of food available, based upon carcass weight of living species, or near relatives, represented by the surviving bone fragments. The weights refer to total food available indicated by the minimum number of individuals that partially survive within the cave.
Body part - the surviving fragments from the cave sediment grouped according to species body part, which indicate the rank order of carcass dismemberment.
Using these criteria, an estimation of biomass, defined by each major ungulate group is tabulated in Figure 116. Each species distribution is described by stratigraphic level. The percentage of survival of skeletal parts is calculated in respect of the number of parts in a live individual. The missing body parts (percentage) is used to calculate the quantity of biomass (meat) that might have been available and used. Surviving skeletal parts fossilised within the cave are also shown.
Figure 116: Reconstruction of body weights and edible food resources based upon the live weights of Minimum Number of Individuals (MNI) and body parts of the major ungulates
Stratigraphic Level
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
Mammoth
Mammoth MNI
0
1
2
0
1
0
7
4
2
2
1
5
3
1
0
1
Survival % MNI
10
6
6
6
2
2
3
2
1
1
1
2
Missing %
90
94
94
94
98
98
97
98
99
99
99
98
edible food (kg)
0
7000
1400
0
7000
0
42000
28000
1400
14000
7000
35000
21000
7000
0
7000
Body Parts
crania
12
4
5
39
10
2
6
8
78
10
3
axial
1
1
3
1
1
limbs
2
2
1
2
2
2
3
26
1
1
Rhino
Rhino MNI
1
2
2
3
2
3
4
2
1
4
2
16
1
1
1
1
1
Survival % MNI
2
0.4
0.8
2
3
3
6
11
17
6
6
2
7
1
0.7
0.3
Missing %
98
99
99
98
97
97
94
89
83
94
94
98
93
1
99
99
Edible food (kg)
3000
6000
6000
9000
6000
9000
12000
6000
3000
12000
6000
48000
3000
1
3000
3000
3000
Body Parts
crania
4
4
14
13
21
40
79
31
34
18
78
12
1
1
axial
3
1
1
3
2
1
1
6
1
1
limbs
2
1
3
4
4
19
15
10
21
13
18
6
1
Horse
Horse MNI
3
2
2
2
2
2
7
4
3
3
2
10
2
2
1
1
Survival % MNI
2
4
6
6
6
11
4
7
5
7
5
8
6
8
1
1
Missing %
98
96
94
94
94
89
96
93
95
93
95
92
94
92
99
99
Edible food (kg)
900
600
600
600
600
600
21000
12000
900
900
600
3000
600
600
300
300
Body Parts
crania
3
8
14
6
13
26
26
30
17
25
12
51
39
5
1
axial
0
2
0
2
6
0
3
0
0
0
1
1
1
1
0
0
limbs
4
3
7
11
1
10
21
10
3
16
3
65
14
1
0
0
Reindeer
Reindeer MNI
2
5
14
17
12
17
9
7
7
6
3
10
6
4
1
1
Survival % MNI
5
3
3
4
12
3
6
9
8
9
8
2
1
6
Missing %
95
97
97
96
88
97
94
91
92
91
92
98
99
94
Edible food (kg)
600
15000
42000
51000
3600
51000
2700
2100
2100
1800
900
3000
1800
1200
300
300
Body Parts
crania
2
1
10
1
1
1
axial
8
12
14
8
1
7
1
3
7
1
18
2
2
1
limbs
1
9
9
2
5
1
15
4
4
7
1
9
3
1
1
Bison
Bison MNI
1
1
1
1
1
1
2
1
1
1
2
1
1
1
0
1
1
Survival % MNI
1
11
15
11
9
8
8
7
3
9
3
9
3
1
1
1
Missing %
99
89
83
89
91
92
92
93
97
91
97
91
97
91
91
91
Edible food (kg)
1300
1300
1300
1300
1300
1300
2600
1300
1300
1300
2600
1300
1300
1300
0
1300
1300
Body Parts
crania
2
1
10
1
1
1
1
axial
8
9
2
5
1
15
4
4
7
1
9
1
1
1
limbs
1
9
12
14
8
1
7
1
3
7
1
18
1
2
1
Stratigraphic Level
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
The principal concern of this analysis is to attempt to identify the relative importance of potential food resources for human groups and carnivores and which are available within the local/regional area surrounding the site. An important observation concerns the differences in biomass between individual species. Both Mammoth and Woolly Rhinoceros may have lived in family groups or singly. Their presence, in terms of season, within the Creswell area is unknown although both species' remains includes juvenile individuals and it is therefore possible that they were summer or late summer visitors. Their large size makes very large quantities of potential food sporadically available. Both species are difficult to hunt and are high risk in terms of danger. With the exception of a few individual sites, there is no clear evidence of death by hunting or carnivore predation. It is much more likely that the adults of these species were scavenged and that hunting was mainly confined to juveniles.
In the case of small ungulates, Horse, Reindeer and Bison, there is evidence that they had higher reproduction rates and lived in large groups or herds, occasionally of considerable size. All three species are highly mobile and seasonally migrate. Evidence from Pin Hole includes foetal and juvenile Reindeer and Bison, which suggest a presence during spring and summer. Evidence from cast antlers suggest that Reindeer may have remained within the area until autumn. Many Horse remains are of older individuals and there is no clear evidence of the season of occupation. For the purposes of this study, it is assumed that Horses are summer occupants. Clearly all three species, especially Reindeer, offer a much larger biomass and potential food resource for humans and carnivores.
Mammoth remains suggest that the species was of considerable importance as an occasional resource. The remains from stratigraphic levels 11-12 and 6-7 in particular appear to represent a resource varying between 21,000 to 48,000kg of meat, and presumably further options of raw materials. Most other stratigraphic levels have at least one individual present. There is clear evidence that they are of interest to Neanderthals (cut-marks) and carnivores (gnawed bones). The species must have offered an occasional opportunistic resource, probably scavenged. It is difficult to envisage that such an opportunistic use led to a development in the predatory skills or hunting techniques needed to kill such a mobile and dangerous species.
In many respects, the evidence for predation of Woolly Mammoth is similar to that of Mammoth. The species seems to have offered a significant resource in stratigraphic level 11 and in levels 3-4, where meat resources vary between 9000-48,000kg. Bones of the species are plentiful, particularly as they are robust, but there has also been a massive amount of bone destruction. As with Mammoth, the species appears to have been a seasonal and opportunistic resource by both Neanderthals and Hyaena.
The analysis of Horse remains is of particular interest. The largest occurrence in terms of food availability is in level 7, which has evidence of up to 21,000kg. This compares with 3000 in level 11 and low quanties in all other levels. It seems that Horse as a prey is an extremely occasional resource (in terms of thousands of years) but on at least two occasions it was a significant resource. In stratigraphic level 10, there is evidence of Neanderthal groups and Hyaena while in level 7 Hyaena presence is insignificant and it is most likely that human groups were the main predator.
Reindeer are known from all stratigraphic levels, where minimum numbers vary considerably. In the lower part of the sequence, many of the remains are post-cranial, including axial parts, and many of these are fractured and gnawed by carnivores. This would seem to indicate that in levels 5-12, the remains are primarily the result of carnivore (probably Hyaena) predation.
There is a very clear and dramatic increase in remains within levels 1-5, particularly between stratigraphic levels 6-5 where the increase is from 2700kg to 51,000kg. Within these levels, the species offers a greater food resource than all of the other species together. The dramatic increase is associated with later Middle Palaeolithic and Upper Palaeolithic use of the site. Large carnivores are in decline at this point and gnawed bone material is probably the result of European Wolf activity.
European Bison remains are present within the whole sequence but in very low numbers and are dominated by limb and axial parts. A great deal of the material is fractured and/or fractured and carnivore gnawed. Its stratigraphic distribution closely follows that of Hyaena and European Wolf. For all species, the quantities present suggest that such resources were occasional and marginal.
9.9 The acquisition of food resources - Neanderthal
Neanderthal food procurement is assumed to be centrally concerned with hunting or scavenging large ungulates that have grazed and converted vegetable matter into a preferred protein diet. Many other localities offer evidence of this practice but there is also a great lack of information concerning the possible use of other resources such as vegetable matter and there is no such evidence available from this site. There is very little direct evidence from Pin Hole, or anywhere else, of Neanderthal capabilities regarding butchery practice, preparation, cooking and preservation of food resources and this is particularly the case for such large species.
To Neanderthal groups, a hunted or scavenged carcass represents a food resource in its entirety and the immediate problems are related to optimising consumption and perhaps conservation of resources while facing competition from a wide range of competitors. These would not only include other large carnivores but a wide range of insects and bacteria capable of rendering the carcass inedible or harmful to health. Such competition operates on an hourly scale and is therefore of immediate concern.
The carcass, whatever its size, represents food in a diverse range of forms. Actualist studies, particularly of carnivores, suggest that initial interest concerns the nutritious and often warm, major organs in the abdomen. This is not a dietary or cultural preference - it is pragmatic in the sense that it is easier to digest protein from warm food as opposed to cold. For Neanderthal groups, the assumed lack of containers to aid transport and potentially conserve food is not a relevant issue. Many visceral remains are encased in strong membranes or skin sacks, which are highly portable and accessible. In view of the time available at a kill site, the fact that these body parts may be warm (and therefore more immediately digestible) makes it likely that they were consumed fresh instantly and on site or nearby. It has already been noted that the associated skeletal parts are usually absent from the cave. It is difficult to cite the lack of evidence of parts as evidence but this does appear to indicate that these highly visible body parts may have been of central interest to humans and carnivores, and have received considerable attention on kill or scavenging sites.
Limb and cranial parts require preparation to warm the food and render easier absorption of the protein. Coincidentally they are often rich resources of potential food and are protected by thick layers of membrane that not only deter fungal infestation but also keep the meat fresh, cool and therefore available for later consumption. This suggests that these parts were of secondary interest at a kill site but were of considerable importance in that they could be transported away from the competition of a kill site to a safer and potentially less competitive and cooler environment, such as a cache or cave. This behaviour has been documented for many carnivore species (Kruuk 1972). The advantage of temporary storage for later consumption of such physically large pieces, momentarily available in abundance, is considerable. This advantage would override the increased energy requirement involved in transportation. Although the argument is circumstantial, a procurement practice of this form goes some way to explain why this site, characterised by its difficult access, contains the remnants of large and heavy body parts that appear to have no other function. Within the entire and numerous collection of skeletal parts from Neanderthal horizons, two have shaft cut-marks. Both are limb fragments (tibias) and are ungnawed by carnivores. These may indicate unusual evidence of attempts at food storage and preparation of the preserved, cool meat and aimed at dissecting the limb prior to warming, perhaps by roasting. There is evidence of at least one burnt Reindeer antler from these stratigraphic levels. The fact that both are tibias, each of a different species, suggests that Neanderthals selected the upper rear leg from large carcasses for ease of transport. In addition, the near-complete Woolly Rhinoceros mandibles known from stratigraphic levels 9 and 11 are undamaged by carnivores, which suggest that they are the result of Neanderthal activity. Many of these species' skeletal parts from the lower stratigraphic levels are crania, particularly mandibular parts. This is a clear indication of the transportation of mandibles (with or without crania) away from kill or scavenging sites. For carnivores, the reasons must relate to competition avoidance, and for Neanderthal groups there is the added advantage of preservation and storage of food resources for later use.
There is no direct evidence of hunting or scavenging activity available from the cave. Actualist studies suggest that this is not necessarily a central issue in that the habitat occupied by modern species or similar species is littered with carcasses of natural fatalities that provide an abundance of carrion (Douglas-Hamilton 1972; 1975, Kühne 1963). It is probable that the Quaternary habitat was of a similar nature. Fatalities are reported as fairly common near water sources, where both species spend significant time in their daily lives. Carcasses are easily located by the attentions of both terrestrial and avian carnivores. In these circumstances, major carnivores are quick to benefit from scavenging carcasses in preference to undertaking the high risk and hazardous activity of stalking and killing such a large-sized prey. Many examples of these species are discovered in such Quaternary contexts.
Many studies often portray Neanderthal groups as hunters who may have scavenged when the opportunity arose. There are few studies that consider how a small carnivore, often-poorly equipped, can stalk, isolate and kill a 7000kg aggressive and mobile ungulate such as Mammoth. Such studies have suggested a number of creative strategies, for instance, driving such animals into ponds or over cliffs to disable them. These studies consistently fail to take account of group defensive behaviour. Modern studies show that such strategies are unrealistic against adults and their family groups. Some comment is available from modern records. Douglas-Hamilton in his profession as a game warden and elephant behaviourist makes several comments upon defensive action by an African Elephant bull who was attacked by armed poachers. In one instance, 34 shots were fired at the African Bull elephant who defended his family by chasing, killing and pulverizing six armed poachers (Douglas-Hamilton 1975).
Carnivore groups such as Hyaena are known to take advantage of the opportunity to kill newborn and young during the spring. It is of some interest that many of the Mammoth and Woolly Rhinoceros remains known from Neanderthal horizons within Pin Hole are juvenile. This may be evidence of a similar practice by Neanderthals during the Quaternary.
During the later phases of the Middle Palaeolithic, from stratigraphic level 8 and above, the quantity of Reindeer remains becomes significant and are associated with a decrease in the remains of Mammoth and Woolly Rhinoceros. Body part diversity also increases, with limb and axial fragments generally outnumbering cranial parts. The presence of both parts indicate a large degree of control at kill or scavenging sites, which ensures recovery of more complete carcasses, some of which have been transported into the cave. Reindeer are much smaller in comparison to previous prey and as such is much more prone to destruction. The transport of axial parts may indicate a continued interest in the major organs and abdominal area but in a species where transport of the whole is more easily achieved. There are multiple ethnographic records of Reindeer predators, including humans, showing an interest in the rumen contents (Forde 1971).
During the Later Middle Palaeolithic (levels 6-7), the frequency of surviving Wild Horse remains increases significantly in a context where destruction (missing parts) remains similar to adjacent levels. These levels contain lithics, Hyaena bones and gnawed bone and unfortunately it is not possible to identify a stratigraphic association between these and the Wild Horse remains. Variation of the remains is distinct and similar to that observed for Woolly Rhinoceros, in that relatively large numbers of cranial bones associated with limb fragments survive. This also suggests that there has been a great deal of control at kill sites where one predator has ensured access to all body parts of the prey. On one occasion (levels 6-7), edible food resources from this species have doubled that provided by Woolly Rhinoceros. Remains of Bison are those of single individuals throughout the whole sequence. Body parts are mainly limb and axial pieces with few cranial remains. The relevance of this pattern of body part survival as an indication of Neanderthal activity is uncertain. The dramatic changes in frequency of edible food evidence and its source species may indicate a change in food procurement systems and diet by Neanderthal groups at Creswell.
In summary, the remains suggest that Neanderthal groups initially relied upon the large ungulates, Woolly Rhinoceros and Mammoth. Palaeontological remains indicate that juvenile individuals were a common prey and that these are at times associated with the remains of large body parts derived from adult animals. The occurrence of juveniles indicates a spring or early summer activity. The evidence also suggests that attempts were made to store food resources. There is no direct evidence to indicate hunting or scavenging practice. Heuristically, it seems probable that scavenging from larger natural fatalities and occasional hunting of juveniles offers an appropriate logistic compromise. Unfortunately, there is little evidence of fire and hearths from the cave but the practice of food storage does suggest that this technology may have been available. The changeover from large prey species (particularly Woolly Rhinoceros) to the smaller species (Reindeer and Wild Horse) in the closing stages of Neanderthal use of the cave is significant and would have been the cause of dramatic change in Neanderthal lifestyle. Interestingly this change is associated with distinct changes in lithic technology from robust quartzite cutting edges to a wider range of cutting edges fabricated in flint. These may have been more appropriate for processing smaller prey species.
9.10 Neanderthals - at the edge of the Quaternary world
A great deal of evidence has been considered concerning Neanderthal use of Pin Hole Cave. One observation is absolutely clear - there isn't much surviving evidence. The low number of stone tools are associated with prolific vertebrate remains where analysis suggests destruction or selection to the extent that the palaeontological evidence, at best, seems to represent 10% or less of the more complete picture. Direct evidence is limited to two cut bones and a number of suggestions for others. This also points to the focus of Neanderthal activity being elsewhere, albeit locally. The direct evidence simply indicates use of two large juvenile ungulates associated with a limited range of tools. The question is, what are Neanderthal groups doing in a site that is several hundred kilometres north of their otherwise known distribution and why are they here? At some points in time, they were at the edge of their known world.
Palaeo-ethological and palaeo-ecological concepts suggest that these groups have moved into an area motivated by their vigality in a random fashion and unaffected by inter-species competition (Putman and Wratten 1984)). They display territoriality in the sense that a low number of sites are known to be specific to this region. Vertebrate population evidence is very specific, is diverse at all times and clearly migrates into the area for breeding in spring and summer. Neanderthal interest in juvenile Mammoth and Woolly Rhinoceros is clear but is very rarely indicated by killing or butchery marks on these species. Similarly, there is also no such evidence from large carnivores other than a tendency to gnaw the bone of juveniles. Under these circumstances it is reasonable to assume that both Neanderthal and carnivores have been motivated to follow migrating herds of large ungulates into their breeding areas. Such areas offer increased opportunities for carcass procurement as ungulate populations are often sexually segregated in such regions, which considerably lowers risk, and are also characterised by increased numbers of natural fatalities, including juveniles. Such conditions offer opportunities for lower risk hunting strategy of juveniles and for opportunistic scavenging of adults. The presence of diverse body parts for juveniles and limb parts for adults within Pin Hole seem to confirm this. The selection and transport of such particular body parts to the cave seem to suggest consumption of axial (major organs) parts at source and storage and preservation strategy for limb parts and heads, which are to some extent protected from decay.
The concentration on Mammoth and Woolly Rhinoceros with the Early Middle Palaeolithic changes and Later Middle Palaeolithic levels see a decrease in both species and an increase in Reindeer, Horse and Bison represented by a range of body parts, suggesting control at kill sites and therefore probably hunted. This is associated with a distinct change in the nature of the lithic assemblage, implying that local typological change in artefacts may be functionally motivated and specifically related to trophic change.
9.11 The acquisition of food resources - Upper Palaeolithic
Evidence for the Upper Palaeolithic is known from stratigraphic levels 1-5. These levels also witness a dramatic, 100% increase in surviving Reindeer remains. The percentage of missing skeletal parts anticipated in a live population continues to be of the order of 90%.
Surviving body parts now include those derived from the whole skeleton. Cranial parts are mostly those of antler fragments to the exclusion of teeth and skull bones. Many of these are cast antlers, which must have been separately collected within the region. Procurement now includes transport and processing of whole carcass remains as edible food and material resources. The relevant stratigraphic levels contain multiple examples of cut skeletal parts and cut antler and cut ivory. The latter material is from Mammoth, a species thought to be locally extinct at this time.
Reindeer individuals are known from studies to be more timid than larger prey and are generally more tolerant of human contact and interference. Many recent and modern human groups who prey on the species maintain a pastoralist-type role with a Reindeer group, which alleviates the unpredictability and expense of energy during hunting. Spring and summer occupation is indicated by the presence of juvenile animals. This essentially exposes human groups to contact with clustered groups of female Reindeer, where the male population has dispersed over a larger region. The combination of the need for stable strategies for food procurement by human groups and the herd breeding behaviour patterns of Reindeer ensure that the development of a close relationship is a certainty. The development of such a relationship over time is very likely to be a significant causal factor in the dramatic increase in activity by both species, which is evident within the cave and its region. Although controversial, the description in this report of an engraved human figure suggests that one of the central images may be a juvenile Reindeer carried by a human figure. The utilisation of other prey species remains a possibility and the indications are that all of the candidates are using the site and area for spring and summer feeding and breeding. This pattern of predation ceases in the most recent stratigraphic levels of the cave for reasons that are unclear.
9.12 Upper Palaeolithic groups - the first settlers
The nature of the evidence associated with the Upper Palaeolithic illustrates dramatic change concerning use of the cave. Many regional and European sites have demonstrated both population increase and diversification in the nature of archaeological evidence. The quantity of remains from the cave is very small in comparison to European sites but indicates substantial change within the context of the cave and activity within the regional habitat. The remains of Hyaena and large ungulates are now of low frequency or absent, probably as a result of regional extinction. There is a distinct appearance of evidence for human groups, Wolf and Reindeer. Skeletal parts of both species are present. The frequent presence of Reindeer cranial parts is now the result of the survival of numerous antlers, many of which have evidence of cut and chop marks. Ivory from extinct species also seems to have been collected as a raw material. Associated with this development, the vertebrate populations increase and diversify dramatically and the remains indicate that many species are spring and summer migrants. The presence of breeding groups of Reindeer, known from the evidence of antlers, is of particular interest in that the population sexually segregates in the spring. Females are known from the cave, suggesting that the breeding area was regionally local to the limestone valleys of the Creswell area and was therefore accessible as prey. It is highly likely human groups had or developed a mutualist relationship with Reindeer populations. This may well have been pastoralist practice and there is some suggestion of this from population distribution, which is regionally clumped suggesting some aspect of movement control.
In such circumstances, the possibility of settlement is dramatically increased. Reindeer are migratory but often spend most months in any one year on the move and large herds can take considerable time to pass through individual areas. The question in a location like Creswell is not one of the degrees of prey presence but is a matter of access, daily abundance and location, all factors that generate the need to evolve methods of resource control such as pastoralist methods. Upper Palaeolithic levels in Pin Hole also contain evidence of many other potential prey species, which include a large variety of birds and fish. There is evidence of carnivore predation of some of these species but the presence indicates resources locally that were available at differing times of year.
Use the navigation bar (top) to go to other sections of this publication
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Internet Archaeology content is preserved for the long term with the Archaeology Data Service (ROR). Help sustain and support open access publication by donating to our Open Access Archaeology Fund.
 Home Summary
Home Summary