A North-Western Habitat: the Paleoethology and Colonisation of a European Peninsula
Rogan D.S. Jenkinson
Cite this as: Jenkinson, R.D.S. 2023 A North-Western Habitat: the Paleoethology and Colonisation of a European Peninsula (a comprehensive analysis of excavations in Pin Hole Cave, Creswell Crags), Internet Archaeology 61 (Monograph 31). https://doi.org/10.11141/ia.61.1
8. Northwestern Habitat, Palaeo-ethology and Colonisation into a European Quaternary Peninsula
There have been three main objectives to this study. The first objective has been to describe the unusual archaeological and palaeontological remains excavated from this locality. This has been undertaken with a view to providing a detailed account of the cave, its excavation and the variation in excavated remains. Past research, particularly by Leslie Armstrong, has rather unfairly attracted considerable criticism. The detailed description and inventories included within this work provide an extensive, accessible data set, which includes sources not previously published and which allow a more comprehensive database to assist future projects and review of this locality.
The second objective of study is to examine the taphonomic context of the finds in the hope that criteria can be recognised that explains their presence within this locality. This is particularly the case for Neanderthal and later human groups, whose presence is only known from lithics and several cut bones. The relationship between this artefactual evidence, the resident large carnivores and the prey or food resources of both groups is crucial for an understanding of events in this cave.
A final aspect of interest is to examine the regional and local context of the locality and particularly to consider why it appears isolated in terms of adjacent relevant finds. The ethology of vertebrate species, which are unusually diverse and frequent, is examined to establish if the context of the locality is important in the ecology of species and the provision of an attractive habitat.
8.1 Heterotrophic elements
Studies by Zimov et al. (1995) have proposed an Ecosystem Hypothesis, which implies that the 'Mammoth Steppe' was not only insensitive to climatic variation but was highly variable and could maintain large populations of graminoids and herbs that were formed through ungulate grazing and trampling and resulted in highly productive grasslands. An important principle of the ecosystem hypothesis is that despite the variation in such a system, Guthrie (1990) has suggested that such systems require at least 10 tons per km² to maintain a herbivore biomass. Recent studies have clearly demonstrated that there is no modern analogue of the Quaternary environment. Consideration of the heterotrophic elements of the Late Quaternary in the UK are summarised here according to topographic variation, the availability of routes for ungulate populations and the accessibility of water.
The Quaternary landscape within the Creswell area seems to offer an undulating landscape with valleys and shelters. To the east was a much larger area of lower lying land, potentially at least 10,000 square kilometres, now under the North Sea and in many ways poorly known. When available throughout the climatic and sea level changes of the Quaternary, this area would have contained a diverse terrain with adequate water and an extensive series of eastward-draining rivers.
To the west and north of the Creswell area, higher, undulating land bisected by valleys, is predominantly of limestone bedrock, and therefore largely waterless. Existing evidence indicates the presence of both humans and animals during the Quaternary but of less importance and more sparse. It also has isolated or clumped distributions, which suggests that it is of marginal or specific use to animal populations. In addition to the essentials of water, the availability of salt (an everyday requirement of animal groups) varies. Adequate supplies of salt exist in many areas of the Benelux countries and can be assumed to be available within the terrain now flooded by the North Sea. Surface and therefore available deposits of salt are present in eastern England from both marine sources and the Permo-Triassic rocks within the eastern and central area. The evidence therefore seems to suggest that the narrow band of higher terrain bordering the lower lying region (Doggerland) may be of considerable interest to both humans and animal populations.
The extensive Magnesian Limestone outcrop forms a north to south trending ridge that runs through eastern England. The eastern outcrop of this formation contains a large number of valleys and gorges and caves that are developed along the eastern area of outcrop and which today offer localised shelter for both plant and animal populations. Many reconstructions relevant to the Quaternary, particularly of plant populations, are derived from cave archaeological sites, and are likely to reflect these localised and sheltered ecosystems rather than provide a more regional image. Studies by Coles (1988) have shown that pollen from cave sites tends to reflect the immediately adjacent environment. Many palaeontological sites have been discovered within fluvial deposits, particularly to the east and south-east of the Creswell region (see Stuart 1982 for summary).
A regional summary of Quaternary conditions must await further research, particularly within the current area of the North Sea. Under these circumstances it does appear that the eastern area of the UK may have offered a corridor to animal populations, which extends into the Midlands. This area is characterised by a large eastern area of relatively low-lying land with extensive fluvial systems and is bordered to the west by a distinct area of upland containing many sheltered valleys and caves developed around river systems and which appear, from existing evidence, to represent a considerable variation in regional conditions. This is the essential element of a hypothesis that points to this region serving as a focal point, accessible by extensive fluvial-based routes, and which provided shelter in the form of varied, non-open, terrain and caves. Such circumstances may well have offered ungulate populations more security and individual protection from predation. For human groups, more realistic hunting strategies would have been possible where the opportunities of ambush tactics and prey selection would have been far greater than those in open terrain.
8.2 Autotrophic elements
Studies of vertebrate distribution and ethology indicate that there are few modern comparisons. Many studies of palaeontological sites have demonstrated that the vertebrate population is of Holarctic type and that individual species distribution have expanded and contracted in response to climatic and environmental fluctuation. It is also clear that the original concept of a 'Mammoth Steppe' does not encompass regional and local diversity of biomass. (Bocherens 2003; Guthrie 1982; 1990; Stuart 1977; Stuart et al. 2002; Jenkinson and Sutherland 1984; Řičánková et al. 2015; Semenov 2014) Within a UK context, the data from Pin Hole Cave suggest considerable vertebrate diversity and the area may have contained Heterotrophic elements of specific regional attraction to a vertebrate population.
8.3 Distribution
A number of authors have noted that animal populations occur in greater numbers within the eastern area of the UK (Stuart 1977; 1982) and these appear to concentrate around major fluvial systems. Despite the possibility of bias in the distribution resulting from differential discovery and research, the general distribution appears contained by environmental factors The evidence suggests that an ecological boundary existed to the west and north within which population has a clumped distribution that becomes random and which indicates an 'edge effect' (Odum and Barrett 1971). The eastern UK distribution therefore seems to represent a distinct and separate ecosystem containing a greater number of species than the areas that surround it and with an ecotone that has low numbers of randomly distributed sites. Populations have temporally fluctuated within the area but it has to be assumed that for most species their behaviours have been the same as those known from studies of populations for current species or related species.
8.4 Population dispersal, vigality and colonisation
The movement of both human and vertebrate population into the western peninsula has obviously been strongly influenced by the presence of natural barriers, hetertrophic resources and the vigality or power of movement of individual species (see Putman and Wratten 1984). Inherent in the suggestion is that the area of clumped distribution indicates dispersal unrestricted by barriers and intra-species competition.
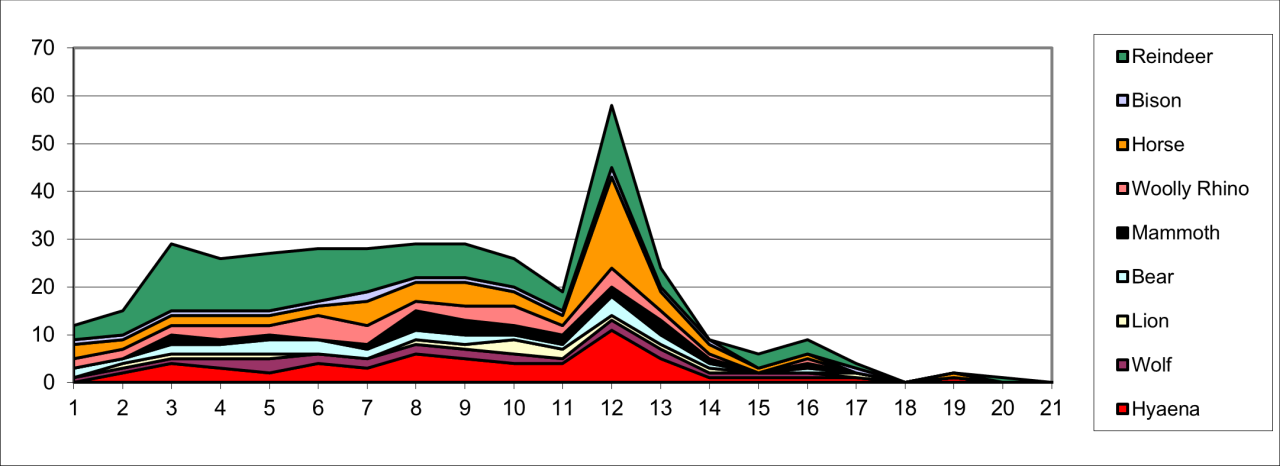
There has been a considerable amount of discussion within the literature concerning the dispersal of both human and animal populations and the causal factors that generated such movement into the UK. In many examples, radiometric dating has suggested a temporal clumping which in turn has suggested periodic dispersal or colonisation. The concept is misleading. The evidence from Pin Hole Cave is regionally rich and diverse, some of the best available for study and is thought to have intermittently accumulated within this cave over a 50,000 year period (Figure 108). In such circumstances, it is heuristically useful to consider the rate and density of such movement. Comparison of the known minimum number of individual animals within such a time frame suggests that temporal density is of very low frequency. The temporal (yearly) occurrence of Hyaena averages one every 1000 years, for Wolf 2000 years, for European Lion 3500 years, for Brown Bear 2000 years, for Mammoth 2000 years, for Woolly Rhinoceros 1500 years, for Wild Horse 1000 years, and for European Bison 3000 years. The nature of this reconstruction is subject to many qualifying factors but it clearly demonstrates the tempo of animal movement and occurrence.
Figure 108: Temporal occurrence of the Minimum Number of Individual animals
Under such circumstances, a concept of 'colonisation' is an overstatement and the vigality and movement are more realistically described as an episodic displacement into the region and where the causal factors are as yet unknown. Figure 108 also illustrates that for many species the Late Quaternary is a period of general long-term decline in frequency, particularly of carnivores, which is associated with a more stable occurrence of Reindeer. The decline within carnivore populations seems to be partially related to decline in large ungulate frequency and is associated with a relative increase in the occurrence of Reindeer and which is increasingly associated with human presence. This indirect evidence may suggest that inter-species competition between increased human activity and decreasing large carnivore density may be due to trophic competition in addition to the inability of large carnivores to adapt to trophic change. There is evidence of a significant disruptive event within stratigraphic levels 11-12 and to some extent in levels 6-7, which coincides with the occurrence of two of the 'slab' layers reported by Armstrong, and which may be related to climatic or geomorphic changes. These levels have revealed an unexplained hiatus within the evidence. The 'peak' in stratigraphic level 12 is demonstrably related to Hyaena residence. The complexities of temporal change evident from the stratigraphic presence of individual animals appears not to support the concept of a distinct and early Mammal Assemblage Zone (MAZ) suggested by Currant and Jacobi (2001) for stratigraphic levels 6-14 and where the widely geographically dispersed locations are considered irrespective of known variation in biome type.
Assessment of dispersion relies on the distribution of both archaeological open-air sites, which are usually known from lithic finds and palaeontological sites known from a range of sedimentary circumstances. There are few known Middle Palaeolithic sites within the regional area surrounding Creswell Crags. Palaeontological sites of both Last Interglacial and Last Glacial age are numerous (see Stuart 1982) and are generally distributed along the eastern margin of the UK.
There seems to be evidence of considerable time gaps in both human and animal population use of the area, particularly in the Late Pleistocene which suggests that a new ecosystem developed within that period. The Later Palaeolithic has a fairly well-established distribution known from the cover sands of South Yorkshire to the north, directly eastwards where rich Later Upper Palaeolithic localities have been discovered at Fardon in Nottinghamshire, and Bradgate Park in Leicestershire There is also considerable evidence increasingly available from Doggerland. Indirect evidence is provided by low numbers of finds of male Reindeer with no females evident at Ravencliffe Cave in Derbyshire (Scott and Hughes 1981), indicating a segregated male group near to the boundary or Edge Effect apparent in the species distribution. Archaeological sites where lithics are often more frequent and diverse than those known from Creswell presently appear to have a random distribution but several are located near major river systems. Both types of site are concentrated within the area of the eastern UK, with occasional localities known from the area to the west.
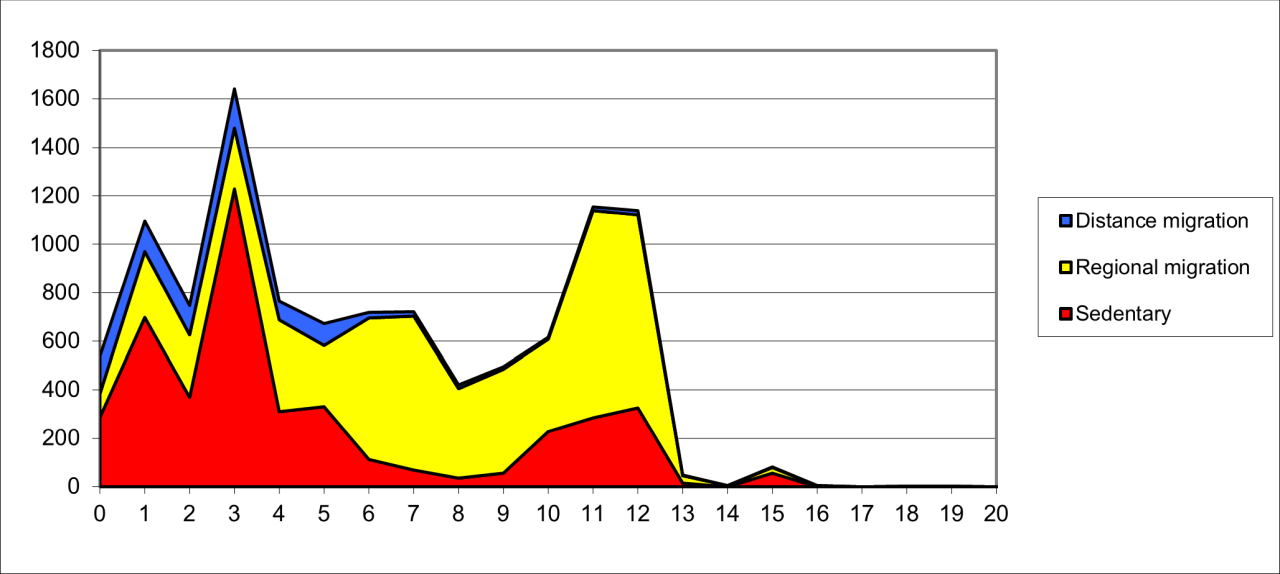
The palaeontological evidence for many species from Creswell Crags, particularly that of Pin Hole Cave, suggests that the population is unusual and of considerable significance in that it is more diverse than the surrounding regional areas. Of the identified species from the cave, 11% are migratory over long distance, 61% migratory over regional distances and 28% semi or fully sedentary and hibernate. This is a pattern of movement that fluctuates temporally but forms a constant feature within the region and is absent from adjoining areas. These are shown in Figure 109.
Figure 109: Population dispersal behaviours of species from Pin Hole
Due to the unreliability of minimum number of individual estimates, bone frequency for all species from each stratigraphic level has been used to construct dispersal patterns and these have been grouped according to the species' modern population dispersal behaviour. Species grouped as regional migrants are those assumed to have moved within the east of England and nearby continental areas. Distance migrants are species, usually bats, birds or fish, that are known to undertake intercontinental migrations.
These data suggest two types of dispersal. Levels 7 to 13 show relatively high frequencies of regionally migrating species, which include large ungulates such as Reindeer, Bison, Woolly Rhinoceros and Mammoth, which are associated with a smaller frequency of sedentary species, including Hyaena, European Brown Bear and Red Fox. Stratigraphic levels 0 to 6 are in sharp contrast, with high frequencies of sedentary species associated with a lower frequency of regionally migrating species and distance migrating species.
It is clear from the analysis that during the early history of the cave occupation was by a low number of carnivores (including humans) associated with a range of migratory ungulates with no evidence of other food resources. The population must therefore have been 'interdependent' upon each other and are likely to have had a similar population dispersal or movement pattern. The uppermost sequence shows a dramatic increase in sedentary species, dominated by small Mustelids, Rodents and European Wolf and these are associated with the appearance of long-range migrants (birds) (Jenkinson and Gilbertson 1984; Jenkinson and Sutherland 1984). These upper levels also show a large increase in human presence, indicated by lithic pieces, and these are associated with Reindeer and Wolf. The high frequency of migrating species results from dispersal motivated by breeding or changes in food procurement. Movement on this scale is typical of ecosystems that experience dramatic seasonal change in temperate conditions and the availability of primary food resources, and these are often found in grassland or steppe regimes populated by large herbivores.
The environmental setting of Pin Hole and similar sites within the area is one of limestone gorges, which border lower lying fluvial and lacustrine systems and may have functioned as refugia for many species. The very low number of known palaeontological localities makes assessment of the dispersion of populations difficult but reliance on actualist behavioural studies enables a number of observations. The presence of long-range migrants, mostly birds, and associated eggs suggest the area was a spring/summer breeding ground in which populations would have been concentrated around water sources. Populations would have been dispersed in clumped sexually segregated groups over very large areas. The potential dispersal and population density is discussed in Reindeer survival. It is clear that direct evidence of their presence has been discovered at other Quaternary localities, which have a large elongated distribution along the eastern UK and the North Sea basin.
Population and dispersal on a continental scale has been the subject of many recent studies. The exhaustive studies by Guthrie (1990) of Alaskan vertebrate faunas have suggested the existence of a Holarctic or steppe fauna characterised by specific species and flora and which has been described as that of a Mammoth Steppe. (Guthrie 1990). Following the original characterisation of this distinct fauna, further studies have described both local variation within the Quaternary Holarctic and the ability of some ungulate populations to develop and sustain such biomes (Killeen et al. 2014). It has become clear that there are several differing elements within the vertebrate population and these have been characterised by Řičánková et al. (2015) as consisting of an arctic population containing: Arctic Fox (Vulpes lagopus), Musk Ox (Praeovibos priscus), a steppe population (Mammoth Steppe Fauna containing the species Mammoth (Mammuthus primigenius), Horse (Equus sp) and European Bison (Bison sp) and Steppe Pika (Spermophilus sp), which tolerated drier but seasonally cold conditions), and these are all associated with a cohabiting population characterised by a preference for more temperate areas and included Hyaena (Crocuta crocuta), European Lion (Panthera leo), Wild Pig (Sus scrofa), Giant Deer or Elk (Megaloceros giganteus or Alces alces), Ibex (Saiga tartarica) and Red Deer (Cervus elaphas). The Quaternary development and expansion of this vertebrate population has been documented throughout the Holarctic region. Řičánková et al. document the expiration (local extinction) of many of these species throughout the Quaternary and a population retreat to the Eurasia steppes, as well as extinction during the Late Glacial and Holocene for many species (Řičánková et al. 2015).
Studies of the megafauna in Pin Hole Cave (Currant and Jacobi 2011) have identified an element of the Mammoth Steppe Fauna, which is also known to be of considerable diversity (Jenkinson 1984, and this study). Studies of the stratigraphic distribution of these species suggest that the Late Quaternary evidence from the cave documents a long period of population decline and expiration for many species prior to the Holocene not apparently related to the expansion of human use of the cave. It is likely that the appearance of more temperate environmental conditions and specifically the afforestation of the region played a significant part in the eastward retreat of a Steppe habitat and its associated species.
8.5 Aggregation
Aggregation of populations (Allee et al. 1949) is apparent within the Creswell region and associated watercourses and there is direct evidence that this is related to breeding. In this sense the locality may have served as a seasonal focus, having sufficient resources for populations to gain security from the dangers of individual predation by maintaining large numbers. Many ungulate species, particularly Reindeer, have a clumped Pleistocene distribution. Similar dispersion, perhaps on a smaller scale can be anticipated for Bison where seasonal requirements for grazing require a large home range. The circumstances for Woolly Rhinoceros and Mammoth are more difficult to assess. Both species have no modern survivor but the evidence from studies of near relatives suggests that they lived in small groups, aggregations of such groups, or were solitary. Mammoth and Horse are thought to have lived in family based groups of unknown size. Both species are known to locally migrate for grazing, water, salt and breeding (Sutcliffe 1985; Walker et al. 1975). Multiple studies have suggested that variation in species dietary preferences and climatic tolerance may be responsible for aggregation in differing biomes.
8.6 Territoriality
Actualism can be inferred using modern analogues which indicate that competition for resources and direct antagonism between species is the prime cause of isolation by groups. For larger sedentary vertebrates, territoriality is often heavily dependent upon food resources, inter-species competition and predation. Within steppe or sub-arctic habitats where access to hetertrophic resources such as plants are of great importance but seasonally can be of restricted access, slow regeneration and of low density, territories are likely to be extensive and exploited by similar species who practice mutualist behaviour. This is often based upon different adaptations or food preferences by species of the same genera and distribution. Wild Horse, Bison and some deer will often coexist in steppe environments but consume different parts of grasses and mosses, with one species eating the taller more robust elements, which exposes the new growth preferred by other species. There is also an element of individual security within large diverse herds. It is probable that ungulates dispersed in smaller sub-groups of the population throughout the area of distribution of water and of their food resources within any one season. Such an area is likely to have generally required vast areas of Eastern England, which were able to seasonally support large numbers of each species. Population concentrations of carnivores, such as Hyaena and Wolves, are known from modern studies to isolate ungulate groups who generally adapt their behaviour to avoid such areas. The regional concentration apparent from the Creswell Caves and Pin Hole Cave may well have produced such an effect in both dispersal and aggregation, thus causing the carnivores themselves to regionally disperse. Indications of such a possibility are available from Conisborough (Anon 1878) where several large carnivore-damaged bones of Woolly Rhinoceros (Coleondonta antiquitatus), Mammoth (Mammuthus primigenius) and Horse (Equus sp) were discovered, from Ravencliffe Cave in Derbyshire (Scott and Hughes 1981), and locally at Langwith Cave (Mullins 1913), Ash Tree Cave and Dead Man's Cave and localities to the south (Jenkinson and Wynne-Griffiths 1986). Small carnivores and rodent species, including Lemmings, have fairly restricted life territories and would have had territories local to the Creswell Gorge where sufficient plants, ground cover and water were accessible.
In his study of the UK Palaeolithic, John Campbell (1977) demonstrated that there are distinct differences between Upper Palaeolithic lithic assemblages from the Creswell area and those known from the Mendip area of the western UK. He suggested that this may be evidence of territoriality with a boundary in Central England. This largely forgotten idea seems to apply to all that is known for both human and vertebrate distribution during the Late Quaternary. Such a concept is in agreement with concepts of pastoralism in the Eastern UK and which is, by definition, territorial.
8.7 Population control - interaction and inter-specific competition
One of the concerns of this study is to comment on autotrophic components of the ecosystem where the palaeontological evidence for these self-feeders is relatively abundant. A useful approach for examining this component is to consider the area in terms of the dynamics of trophic systems, at least for the species that are evident from direct evidence. The question translates into a need to identify the resources that might have been available for predators, including humans, and their necessary resources, which allowed occupation in such an apparently peripheral area of the Quaternary world. Previous studies have attempted such an approach, particularly for studies of human groups (Campbell 1977), but many of these studies have been based upon ethnographic data and have largely been concerned with small and restricted territorial areas centred upon a main base and essentially a semi-sedentary life style for both humans and animal populations. There is little circumstantial ecological evidence that offers support to any of these views. Evidence for inter-species competition and predation is apparent from analysis of the similarities in distribution of some groups of the vertebrate population.
Figure 110: Stratigraphic distributions of small carnivores raptor birds and rodents, birds and fish
The distribution of small carnivores consistently includes Red Fox (Vulpes vulpes) and Badger (Meles meles), and within the uppermost levels of the cave, small Mustelid species appear episodically. These levels also contain small numbers of raptors, which include several species of Eagle, Hawk and Owl. Many of these species are known from actualist studies to predate on rodents, birds and fish by means of direct hunting or by scavenging. Comparison of their stratigraphic distribution within the cave shows that they have had similar distribution during the cave's history (Figure 110). The presence of specific indications of small carnivore predation in the form of bite marks has already been noted for species of Fish, Grouse, Ducks and Geese. Long-term variations in prey frequency are not replicated by predatory species but are associated with relative stability of small carnivore populations, which may suggest that predators were adaptive and responded to prey availability and variation. The distribution of Wolf (Canis lupus), shown in Figure 32 is also remarkably similar to that of rodents (Figure 86) There is a clear increase in bird frequency that is associated with the appearance of Raptors within the upper levels of the cave. The significance of the major depressions in frequency within levels 5-6 and 8-9 is unknown. Both depressions occur within the levels where Armstrong noted the existence of slab layers consisting of limestone clast and may be due to geomorphic events affecting stratigraphic distribution, rather than change within inter-species relationships.
Direct competition in the form of predation is clearly indicated by fracture, gnawing, biting, surface licking and lithic cut-marks upon the remains of prey species. Competition for resources is apparent in that evidence of the earliest Middle Palaeolithic contains cut-marked bone of both Mammoth and Woolly Rhinoceros. The same levels contain carnivore-damaged bone of the same species. The combination of Middle Palaeolithic tools, ungnawed cut bones, fractured bone and Hyaena gnawed bone within the same 'stratigraphic levels' offers comment on the limited degree of temporal definition available from this site but also clearly demonstrates that human use of the cave was not contemporary with Hyaena residence.
Figure 111: Stratigraphic distribution of human groups and the major large ungulate species
The distribution of human groups indicated by artefact frequency and the minimum numbers of the ungulate species Reindeer (Rangifer tarandus) and Horse (Equus sp) is striking. An inter-species relationship is confirmed by direct evidence in the form of cut and incised bones of some of these species. (Figure 111). The frequency increase of human groups associated with relative stability in Reindeer numbers suggest an increased proficiency in predation by human groups upon the species and there is evidence of the use of its body parts (antler) as resources. The relationship with Horse appears significant within the lower levels, associated with Middle Palaeolithic artefacts, but diminishes in the longer term. Both Mammoth (Mammuthus primigenius) and Woolly Rhinoceros (Coleondonta antiquitatis) appear to be marginal resources for human groups but are known to have been utilised as a resource for ivory and incised bone, which may be the product of skinning activity. The increase of ivory and bone working on the bone of apparently extinct species may indicate the collection of sub-fossil material, particularly during the Later Palaeolithic where there is evidence for the collection of Jurassic fossils and marine mollusca.
There has been much discussion concerning the relevance of Bison (Bison sp) as a human resource and the development or use of specific artefacts (White and Pettitt 2011) appropriate for butchering their robust carcasses. The distribution shown in Figure 111 suggests that they were of marginal interest to early Neanderthal groups and carnivores, which is perhaps not surprising given their size, mobility and potential for aggressive defensive behaviour when confronted by predators. The evidence also shows that they were of considerable significance during later periods of Neanderthal occupation.
These levels also contain the contemporary remains of Wolf (whose main prey may have been Reindeer and Rodents) and which in turn suggests that human groups, Hyaena and Wolf had competitive relationships. The available evidence is not of sufficient detail to distinguish events, episodes or temporally isolated occupations, but its constancy over time suggests competition. It is interesting to note that the low frequency of Wolf in early stratigraphic levels dramatically changes with the demise of Hyaena populations in a context where all levels contain high frequency of fractured and gnawed vertebrate bone and where Wolf must have been the main causal factor. Many of the associated unidentified vertebrate bone fragments have recently been identified using DNA fingerprinting (Buckley et al. 2017) to the common prey Reindeer. The significant rise in numbers of Wolf and the increased human use of the cave characterises levels 1 to 5 and is associated with large numbers of Reindeer with samples of both cut-marked and worked bone, which are occasionally associated with gnawed and cast Reindeer antler. There is no evidence of episodic variation in species presence or activity. Wolf is present in consistently low individual numbers of mixed age and must represent family groups rather than large packs. This also suggests the possibility that the relationship between human groups and Wolf might have been one of mutualism, with both having access to a common prey with no apparent detrimental effect on themselves and in fact in circumstances where the density of both species increases. Both species cease to occupy the cave at the same time (Figure 112). The nature of the data inhibits further examination of the relationship but it is distinctly possible that it was cooperative.
Figure 112: Inter-species relationship between Humans, Wolf and Reindeer
A great deal of evidence is available that indicates a state of mutualism between species, notably for large ungulates. These species are often adapted to consume different components of the same grass or moss genera. This method of specific species adaptation not only maximises resources but is also likely to give rise to greater numbers of each species using the same food resource in a context where a diversity of species occurring in greater numbers offers some individual protection against carnivores.
8.8 Parasitism
Parasitism tends to be quantitatively of small occurrence in whole populations but has a very negative effect upon individuals. This form of interaction is common in many species but in this context is of considerable importance for ungulates. Both Artiodactyla and Perissodactyla experience seasonal parasitism, particularly in warm lower altitude landscapes with abundant water resources. Modern examples are known for many ungulate species (Forchhammer et al. 2001; Mooring and Hart 1992; Helle et al. 1992; Duncan and Vigne 1979). Intra-species competition and parasitism would have been of considerable significance for ungulate populations and their behavioural response, movement, regional aggregation, social structure and susceptibility to predation by carnivores and human groups. These behaviours are of particular relevance for such populations living within the varied and particular habitats offered by the gorges and valleys of the Creswell region.
Many ungulate groups are attacked during the spring and summer months by Diptera insects which include Mosquitoes, Black Flies, Biting Midges, Nose Bot Fly and Warble Fly. It is not certain but it is likely that parasites similarly affected both Mammoth and Woolly Rhinoceros. Many parasite species lay eggs in the nasal area or rectum of the host and this leads to the development of cardiovascular and movement difficulties. Modern studies have shown that some species suffer up to 200 attacks at any one time (Mooring and Hart 1992).
Host species have a number of behaviours that deter parasites, which include speed and style of individual movement (Reindeer), shaking and tail movement (Horses and Bovids). Host species commonly regionally migrate to higher, cooler areas, particularly during the breeding period, in an attempt to reduce infestation. Parasitic species are also able to move rapidly over large distances and locate hosts by smell but they are deterred by host movement, cooler temperatures and higher altitude. Damage and destruction of areas of the skin and pelt are often common within infested individuals, which for human groups, who themselves experience similar problems, makes pelts useless as a raw material. Such negative interaction can also serve as a motivation not only for movement but can isolate populations within particular areas of their home range. This was probably of some importance for populations attempting to spend summer in the low-lying hills of Doggerland. It is possible that recent studies, which have drawn attention to the concentrations of human settlement in valleys and lower lying areas and the nutritional value to ungulates of these areas (Brown et al. 2013), may be documenting winter feeding areas for both ungulate and human populations.
There are a number of additional behaviours related to ungulate groups that give some protection against carnivores, which include the practice of local aggregation, This was first noted by Hamilton (1971), who coined the term 'selfish herd' to describe the practice of individual ungulates moving further into the herd and forming more dense groups as a means of diluting the individual risk of predation, particularly by parasites (Mooring and Hart 1992). This behaviour offered parasites greater choice of host but also allowed individual ungulates greater protection, security and shelter. It also offered other larger carnivores, including humans, a greater opportunity for predation. Herd movement designed to reduce parasitism often results in herds moving into sheltered foliated valleys where conditions might have been cooler, darker and windy in a very local sense. This behavioural pattern must have been of considerable significance within the Creswell area which is characterised by such topographic features. While offering protection and shelter to both ungulate populations, human groups and primary predators, this also locally concentrates both predator and prey into areas where movement is more restricted offering predators more opportunity for the concealment and ambush tactics necessary to hunt a more concentrated but highly mobile ungulate species.
There is no currently available direct evidence of parasitic species from Pin Hole Cave, but indirect evidence from studies of modern species populations suggests that the interaction was probably just as important in the past as it is today. Studies by Turner (1981) have suggested that many of the Horses known from Pin Hole are older females and studies for this report have demonstrated a relatively large presence of foetal and juvenile individuals. It could be the case that the reduced powers of movement of this normally highly mobile and potentially aggressive species results from both age, parasitism and pregnancy has increased the likelihood of predation by carnivores and therefore accumulation within the cave. The specific interaction between parasites and ungulate populations is also likely to have been a significant motivation for regional migratory movement and dispersal.
Use the navigation bar (top) to go to other sections of this publication
Internet Archaeology is an open access journal based in the Department of Archaeology, University of York. Except where otherwise noted, content from this work may be used under the terms of the Creative Commons Attribution 3.0 (CC BY) Unported licence, which permits unrestricted use, distribution, and reproduction in any medium, provided that attribution to the author(s), the title of the work, the Internet Archaeology journal and the relevant URL/DOI are given.
Internet Archaeology content is preserved for the long term with the Archaeology Data Service (ROR). Help sustain and support open access publication by donating to our Open Access Archaeology Fund.
 Home Summary
Home Summary



{kind=link}